Applications to Sports Performance with a Focus on Intermittent High-Intensity Exercise
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
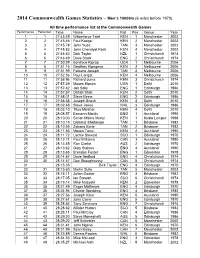
2014 Commonwealth Games Statistics – Men's
2014 Commonwealth Games Statistics – Men’s 10000m (6 miles before 1970) All time performance list at the Commonwealth Games Performance Performer Time Name Nat Pos Venue Year 1 1 27:45.39 Wilberforce Talel KEN 1 Manchester 2002 2 2 27:45.46 Paul Kosgei KEN 2 Manchester 2002 3 3 27:45.78 John Yuda TAN 3 Manchester 2002 4 4 27:45.83 John Cheruiyot Korir KEN 4 Manchester 2002 5 5 27:46.40 Dick Taylor NZL 1 Christchurch 1974 6 6 27:48.49 Dave Black ENG 2 Christchurch 1974 7 7 27:50.99 Boniface Kiprop UGA 1 Melbourne 2006 8 8 27:51.16 Geoffrey Kipngeno KEN 2 Melbourne 2006 9 9 27:51.99 Fabiano Joseph TAN 3 Melbourne 2006 10 10 27:52.36 Paul Langat KEN 4 Melbourne 2006 11 11 27:56.96 Richard Juma KEN 3 Christchurch 1974 12 12 27:57.39 Moses Kipsiro UGA 1 Delhi 2010 13 13 27:57.42 Jon Solly ENG 1 Edinburgh 1986 14 14 27:57.57 Daniel Salel KEN 2 Delhi 2010 15 15 27:58.01 Steve Binns ENG 2 Edinburgh 1986 16 16 27:58.58 Joseph Birech KEN 3 Delhi 2010 17 17 28:02.48 Steve Jones WAL 3 Edinburgh 1986 18 18 28:03.10 Titus Mbishei KEN 4 Delhi 2010 19 19 28:08.57 Eamonn Martin ENG 1 Auckland 1990 20 20 28:10.00 Simon Maina Munyi KEN 1 Kuala Lumpur 1998 21 21 28:10.15 Gidamis Shahanga TAN 1 Brisbane 1982 22 22 28:10.55 Zakaria Barie TAN 2 Brisbane 1982 23 23 28:11.56 Moses Tanui KEN 2 Auckland 1990 24 24 28:11.72 Lachie Stewart SCO 1 Edinburgh 1970 25 25 28:12.71 Paul Williams CAN 3 Auckland 1990 25 26 28:13.45 Ron Clarke AUS 2 Edinburgh 1970 27 27 28:13.62 Gary Staines ENG 4 Auckland 1990 28 28 28:13.65 Brendan Foster ENG 1 Edmonton 1978 29 29 28:14.67 -

GRTC to Induct Olympian John Tuttle and Community Leader/Runner David Bischoff Into Hall of Fame
Media Release: January 17, 2007 Contact: Michael Reif (585-377-1041 or email [email protected]) Or GRTC President Tom Perry, email: ([email protected]) For more information on GRTC Hall of Fame see: http://www.grtconline.org/) GRTC to induct Olympian John Tuttle and Community Leader/Runner David Bischoff into Hall of Fame The Greater Rochester Track Club (GRTC) is pleased to announce the selection of Olympian John Tuttle of Douglassville, Georgia and Runner/Community leader David Bischoff, of Fairport, NY to the GRTC Hall of Fame. Both will be inducted at the GRTC’s Annual Banquet on Saturday January 27, 2007 at the Diplomat Banquet Center 1956 Lyell Avenue, Rochester from 5-9 PM. The dinner is open to the public however reservations are to be made by January 22 to Pete Van Peursem at 34 Denishire Drive, Rochester, NY 14624 (dinner tickets are $17 per person in advance, $22 at the door) David Bischoff of Fairport is an outstanding multi-sports athlete and runner. He is the team captain of his Genesee Valley Harriers (GVH) Running Club Master’s team and is a member of their National Championship Cross Country team (2005 @5K) as well as several runner-up national championship teams. He led the USA Masters Men to victory over Canada at this year’s annual CanAm Master’s Cross Country Challenge at Watkins Glen, NY. His running and other athletic accomplishments are numerous. In addition to David’s athletic prowess in multiple sports he is being recognized for his contributions as a leader in the sports community through his promotion and dedicated volunteer work in major endeavors including youth sports development. -

Paul Tergat O Most Observers It Came As No Great Surprise Tergat: Thank You Very That Paul Tergat Produced a WR 2:04:55 in Much
T&FN INTERVIEW Paul Tergat o most observers it came as no great surprise Tergat: Thank you very that Paul Tergat produced a WR 2:04:55 in much. I knew that I had T Berlin. So great are the talents of this legendary the potential. I knew that I by Sean Hartnett Kenyan—be it on the track, harrier course or the had the ability for bringing roads—that he faced WR expectations in every one down the World Record for of his previous five marathons. Tergat steadfastly the marathon, maybe by a maintained that “the marathon is a completely few seconds. But it was a big different event and I have much to learn.” surprise for me to go under This says much about the 34-year-old Kenyan, 2:05. Whatever you have whose quest for running greatness is matched by been putting in—in terms his passion for knowledge on all fronts. When he of energy, in terms of mental is not training his days are filled with a multitude preparedness and physical of family, business, and charitable activities, all torture—it is sweet when you the while juggling a couple of active cell phones. have such great moments. Conversation with Tergat ranges easily from world T&FN: Many people pre- issues to athletics or his homeland, and is always dicted that you would be the spiced with a bit of humor. WR holder right off the bat. But While Tergat is the epitome of a Kenyan distance the marathon is a very difficult runner, he is far from typical and did not even event, and you have made slow begin his running career until he completed his step-by-step progress. -

The Runner's Rite of Spring®
The Runner’s Rite of Spring® 1995 (April 9) women’s title in 54:28. A total of $35,000 was given to Chil- Pre-race speculation centered on the Kenyans cracking the dren’s Hospital, bringing the total charity contribution to 46:00 barrier for the first time. With world cross country over $140,000. runner up Ismael Kirui and 1994’s third place finisher Jos- phat Machuka as the top seeds, the stage was set (Sigei was 1998 (April 5) out due to injury). Race day dawned with no wind, although A new course featuring an out-and-back section on Rock temperatures were in the upper 50s and the humidity was Creek Parkway delighted the 5,800 finishers. South Afri- high. As expected, Kirui and Machuka went right to work, can Colleen DeReuck proved that the layout was fast as dropping further below world record pace with each pass- she shattered Rose Cheruiyot’s World Record for 10 Miles ing mile. Suffering from blisters, Machuka dropped off by of 51:39 — set in the 1995 race —with a scintillating 51:16 5 miles. Kirui slowed to a 4:41 ninth mile, but responded performance that placed her 23rd overall. (She would have with a 4:31 closer to smash Sigei’s year-old mark with a been the overall winner of the 1973 race!)DeReuck finished 45:38. Just over six minutes later Rose Cheruiyot of Kenya, over two minutes ahead of runner-up Marian Sutton of who had set a world 5K record a week before, collected the Great Britain. -

Past Award Recipients
Award Year Recipient Robert Barnes Distinguished Service 1988 Robert Weiss, DPM(1st award recipient) 1989 Richard Schuster, DPM 1990 Rob Roy McGregor, DPM 1992 Justin Wernick, DPM 1993 John Weed Family 1994 Thomas Sgarlato, DPM 1995 Thomas Amberry, DPM 2000 William Van Pelt, DPM4 2004 Lloyd Smith, DPM 2005 David Davidson, DPM 2006 Patrick Nunan, DPM 2009 Jeffrey Ross, DPM 2010 Richard Bouche, DPM 2011 Timothy Dutra, DPM 2014 Matthew Werd, DPM 2015 John McNerney, DPM 2016 Stanley James, MD 2017 Michael Chin, DPM President's Award 2008 Mark Wagner, DDS 2015 Arthur Gudeon, DPM 2017 Robert Anderson, MD 2018 Mark Cucuzzella, MD Richard Schuster, DPM 1993 Kevin Kirby, DPM 1995 Richard Blake, DPM 2005 Neil Humble, DPM 2008 Richard Bouche, DPM 2009 Simon Bartold 2010 Stephen Pribut, DPM 2011 Amol Saxena, DPM Outstanding Service Award 2008 William Van Pelt, DPM 2009 Robert Frimmel, DPM 2010 Brooks Sports - Mike Billish 2015 David Levesque, DPM George Sheehan Memorial Award 1994 Michael Burns, DPM 1995 Steven Subotnick, DPM Award of Excellence 2006 Mary Moore - Ireland Lifetime Achievement Award 2018 Richard Gilbert, DPM Robert Barnes Legecy Award 2006 Harry Hlavac, DPM John Pagliano Golden Foot Award 1983 Terry Fox (In Memory of) 1984 George Sheehan, MD 1985 Ron Culp, Athletic Trainer, Miami Heat 1986 F. Story Musgrave, Spacewalker 1987 Walter Peyton, Former Chicago Bear 1987 Doug Collins, Former Chicago Bulls Coach 1988 Bill Rodgers, Distance Runner 1991 Mary Decker Slaney, Track Star 1992 George Bush, Former US President 1992 Warren Moon, Former Houston Oiler 1993 James Garrick 1996 Jim Ryun, Distance Runner 1997 Kurt Marsh, Former Oakland Raider 2000 Mayor John Street (Philadelphia) 2006 Tom Amberry, DPM 2006 Governor Michael Huckabee 2007 Amby Burfoot, Editor, Runners World 2008 David Beckham, LA Galaxy Soccer 2009 Khalid Khannouchi, Marathon Runner 2010 Alberto Salzar 2014 Brooks Johnson 2014 Paula Ratcliff 2016 Nike Oregon Project 2017 Nathan Reitzehein 2018 Gary Hall, Jr. -

IMPRESSIONEN IMPRESSIONS Bmw-Berlin-Marathon.Com
IMPRESSIONEN IMPRESSIONS bmw-berlin-marathon.com 2:01:39 DIE ZUKUNFT IST JETZT. DER BMW i3s. BMW i3s (94 Ah) mit reinem Elektroantrieb BMW eDrive: Stromverbrauch in kWh/100 km (kombiniert): 14,3; CO2-Emissionen in g/km (kombiniert): 0; Kraftstoffverbrauch in l/100 km (kombiniert): 0. Die offi ziellen Angaben zu Kraftstoffverbrauch, CO2-Emissionen und Stromverbrauch wurden nach dem vorgeschriebenen Messverfahren VO (EU) 715/2007 in der jeweils geltenden Fassung ermittelt. Bei Freude am Fahren diesem Fahrzeug können für die Bemessung von Steuern und anderen fahrzeugbezogenen Abgaben, die (auch) auf den CO2-Ausstoß abstellen, andere als die hier angegebenen Werte gelten. Abbildung zeigt Sonderausstattungen. 7617 BMW i3s Sportmarketing AZ 420x297 Ergebnisheft 20180916.indd Alle Seiten 18.07.18 15:37 DIE ZUKUNFT IST JETZT. DER BMW i3s. BMW i3s (94 Ah) mit reinem Elektroantrieb BMW eDrive: Stromverbrauch in kWh/100 km (kombiniert): 14,3; CO2-Emissionen in g/km (kombiniert): 0; Kraftstoffverbrauch in l/100 km (kombiniert): 0. Die offi ziellen Angaben zu Kraftstoffverbrauch, CO2-Emissionen und Stromverbrauch wurden nach dem vorgeschriebenen Messverfahren VO (EU) 715/2007 in der jeweils geltenden Fassung ermittelt. Bei Freude am Fahren diesem Fahrzeug können für die Bemessung von Steuern und anderen fahrzeugbezogenen Abgaben, die (auch) auf den CO2-Ausstoß abstellen, andere als die hier angegebenen Werte gelten. Abbildung zeigt Sonderausstattungen. 7617 BMW i3s Sportmarketing AZ 420x297 Ergebnisheft 20180916.indd Alle Seiten 18.07.18 15:37 AOK Nordost. Beim Sport dabei. Nutzen Sie Ihre individuellen Vorteile: Bis zu 385 Euro für Fitness, Sport und Vorsorge. Bis zu 150 Euro für eine sportmedizinische Untersuchung. -

Ataconic Running Life S
Ataconic running life S ^jj^^^g^www.runner.org• T • R • R November• C 1999* ^ RUNNING INTO 2000 NICK JUBOK GIVING WATER TO GIA BOULOS IN THE LAST LEG OFTHE REACH THE BEACH RELAY C A N W E S U R V I V E Y 2 K ? M A R K Y O U C A L E N D A R S N O W ! 12/4/99 10:00 AM X-Country Relays P e t e r M e i s l e r 265-9674 12/5/99 11:00 AM Couples Relay Steve Calidonna 736-7607 2/6/00 10:00 AM F r e e z e r 5 K Jerry Radlauer 764-9056 Barbara Brittan 747-7431 2/20/00 10:00 AM F r e e z e r 5 M i l e r Jerry Radlauer 764-9056 Barbara Brittan 747-7431 The Taconic Road Runners Club Presents the «i<!ross Country Relay Blue Mountain Reservation Saturday December 4,1999 Starting time 10:00 A.M. Female-3 ninners per team Fee $15/team Mate^ numers per team Fee$20/team All nmners con^pete on the same course, which is 3+ miles and changes every year. The race begins and ends at the Bhie Mountain Lodge. It is a good idea not to wear your newest shoes for this event. There will be a pancake breakfest after the race and showws are available at the Lodge. Awards for male ^d female teams in the following categories: High School, Open and Master. -

Run with a Friend... May 2000 Presidential Message April 2000 B.A.A
Run with a friend... www.mainetrackclub.com May 2000 Presidential Message April 2000 B.A.A. BOSTON Dear MTC Members John Hancock Sports and Fitness Expo Thank you to all who turned out to help with the Boys and Giils club 5 Mile Road Race. It takes alot of volunteers for this race, Thank you for being there. Once again it was another interesting race (Boys Club). I have a thought "from now on, no Police car leads a race unless someone who knows the course is riding with them"! As for the race starting at the drop of the hat and the cannon going off 2 seconds later, well there is no help for that! The And because we couldn't get the times to dump, I have been practicing ever since Howard Spear has shown me the error of my ways, I now know to Champions use that RESETBUTTON!Jean Thomasand MaryAnneChampeonsaved the day with their fast fingers and teamwork to input Bib Numbers and Trophy Times. Bob Aube says the chute went well (overall it did) and we got back the two tabs that blew away thanks to Ann Strout and Mike Reali's dad (Amedeo). I did get one phone call from a runner who wasn't listed in the The first permanent paper. I was able to find him on the back up sheet so I cold give him his Boston Marathon time (his tab could still be blowing in the wind!). Our apologies to the two trophy, it will showcase runners who came after we took the chute the next 100 years of down. -

Course Records Course Records
Course records Course records ....................................................................................................................................................................................202 Course record split times .............................................................................................................................................................203 Course record progressions ........................................................................................................................................................204 Margins of victory .............................................................................................................................................................................206 Fastest finishers by place .............................................................................................................................................................208 Closest finishes ..................................................................................................................................................................................209 Fastest cumulative races ..............................................................................................................................................................210 World, national and American records set in Chicago ................................................................................................211 Top 10 American performances in Chicago .....................................................................................................................213 -

Table of Contents
Media Table of contents Media information & fast facts ......................................................................................................... 3 Important media information ....................................................................................................................................................4 Race week Media Center..............................................................................................................................................................4 Race week schedule of events ..................................................................................................................................................7 Quick Facts ...........................................................................................................................................................................................8 Top storylines ......................................................................................................................................................................................10 Prize purse .............................................................................................................................................................................................13 Time bonuses ......................................................................................................................................................................................14 Participant demographics ............................................................................................................................................................15 -

6 World-Marathon-Majors1.Pdf
Table of contents World Marathon Majors World Marathon Majors: how it works ...............................................................................................................208 Scoring system .................................................................................................................................................................210 Series champions ............................................................................................................................................................211 Series schedule ................................................................................................................................................................213 2012-2013 Series results ..........................................................................................................................................214 2012-2013 Men’s leaderboard ...............................................................................................................................217 2012-2013 Women’s leaderboard ........................................................................................................................220 2013-2014 Men’s leaderboard ...............................................................................................................................223 2013-2014 Women’s leaderboard ........................................................................................................................225 Event histories ..................................................................................................................................................................227 -
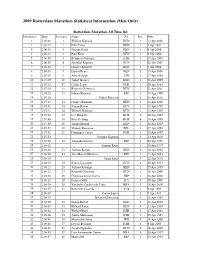
2009 Rotterdam Marathon Statistical Information (Men Only)
2009 Rotterdam Marathon Statistical Information (Men Only) Rotterdam Marathon All Time list Performances Time Performers Name Nat Place Date 1 2:05:49 1 William Kipsang KEN 1 13 Apr 2008 2 2:06:14 2 Felix Limo KEN 1 4 Apr 2004 3 2:06:38 3 Sammy Korir KEN 1 9 Apr 2006 4 2:06:44 4 Paul Kirui KEN 2 9 Apr 2006 5 2:06:50 5 Belayneh Dinsamo ETH 1 17 Apr 1988 6 2:06:50 6 Josephat Kiprono KEN 1 22 Apr 2001 7 2:06:52 7 Charles Kibiwott KEN 3 9 Apr 2006 8 2:06:58 8 Daniel Rono KEN 2 13 Apr 2008 9 2:07:07 9 Ahmed Salah DJI 2 17 Apr 1988 10 2:07:09 10 Japhet Kosgei KEN 1 18 Apr 1999 11 2:07:12 11 Carlos Lopes POR 1 20 Apr 1985 12 2:07:18 12 Kenneth Cheruiyot KEN 2 22 Apr 2001 13 2:07:23 13 Fabian Roncero ESP 2 18 Apr 1999 14 2:07:26 Fabian Roncero 1 19 Apr 1998 15 2:07:33 14 Charles Kamathi KEN 3 13 Apr 2008 16 2:07:41 15 Simon Biwott KEN 3 18 Apr 1999 17 2:07:42 16 William Kiplagat KEN 1 13 Apr 2003 18 2:07:44 17 Lee Bong-Ju KOR 2 19 Apr 1998 19 2:07:49 18 Kim Yi-Yong KOR 4 18 Apr 1999 20 2:07:50 19 Jimmy Muindi KEN 1 10 Apr 2005 21 2:07:51 20 Vincent Rousseau BEL 1 17 Apr 1994 22 2:07:51 21 Domingos Castro POR 1 20 Apr 1997 23 2:07:53 Josephat Kiprono 2 13 Apr 2003 24 2:07:54 22 Alejandro Gomez ESP 2 20 Apr 1997 25 2:08:02 Sammy Korir 3 20 Apr 1997 26 2:08:02 23 Jackson Koech KEN 2 10 Apr 2005 27 2:08:09 24 Jose Manuel Martinez ESP 3 13 Apr 2003 28 2:08:14 Samy Korir 3 22 Apr 2001 29 2:08:19 25 Simon Loywapet KEN 4 20 Apr 1997 30 2:08:21 26 Joshua Chelanga KEN 1 15 Apr 2007 31 2:08:22 27 Kenneth Cheruiyot KEN 1 16 Apr 2000 32 2:08:30 28 Francisco