(1 → 4)-Β-D-Glucan Is a Component of Cell Walls in Brown Algae
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

New Zealand's Genetic Diversity
1.13 NEW ZEALAND’S GENETIC DIVERSITY NEW ZEALAND’S GENETIC DIVERSITY Dennis P. Gordon National Institute of Water and Atmospheric Research, Private Bag 14901, Kilbirnie, Wellington 6022, New Zealand ABSTRACT: The known genetic diversity represented by the New Zealand biota is reviewed and summarised, largely based on a recently published New Zealand inventory of biodiversity. All kingdoms and eukaryote phyla are covered, updated to refl ect the latest phylogenetic view of Eukaryota. The total known biota comprises a nominal 57 406 species (c. 48 640 described). Subtraction of the 4889 naturalised-alien species gives a biota of 52 517 native species. A minimum (the status of a number of the unnamed species is uncertain) of 27 380 (52%) of these species are endemic (cf. 26% for Fungi, 38% for all marine species, 46% for marine Animalia, 68% for all Animalia, 78% for vascular plants and 91% for terrestrial Animalia). In passing, examples are given both of the roles of the major taxa in providing ecosystem services and of the use of genetic resources in the New Zealand economy. Key words: Animalia, Chromista, freshwater, Fungi, genetic diversity, marine, New Zealand, Prokaryota, Protozoa, terrestrial. INTRODUCTION Article 10b of the CBD calls for signatories to ‘Adopt The original brief for this chapter was to review New Zealand’s measures relating to the use of biological resources [i.e. genetic genetic resources. The OECD defi nition of genetic resources resources] to avoid or minimize adverse impacts on biological is ‘genetic material of plants, animals or micro-organisms of diversity [e.g. genetic diversity]’ (my parentheses). -

The Taxonomy and Biology of Phytophthora and Pythium
Journal of Bacteriology & Mycology: Open Access Review Article Open Access The taxonomy and biology of Phytophthora and Pythium Abstract Volume 6 Issue 1 - 2018 The genera Phytophthora and Pythium include many economically important species Hon H Ho which have been placed in Kingdom Chromista or Kingdom Straminipila, distinct from Department of Biology, State University of New York, USA Kingdom Fungi. Their taxonomic problems, basic biology and economic importance have been reviewed. Morphologically, both genera are very similar in having coenocytic, hyaline Correspondence: Hon H Ho, Professor of Biology, State and freely branching mycelia, oogonia with usually single oospores but the definitive University of New York, New Paltz, NY 12561, USA, differentiation between them lies in the mode of zoospore differentiation and discharge. Email [email protected] In Phytophthora, the zoospores are differentiated within the sporangium proper and when mature, released in an evanescent vesicle at the sporangial apex, whereas in Pythium, the Received: January 23, 2018 | Published: February 12, 2018 protoplast of a sporangium is transferred usually through an exit tube to a thin vesicle outside the sporangium where zoospores are differentiated and released upon the rupture of the vesicle. Many species of Phytophthora are destructive pathogens of especially dicotyledonous woody trees, shrubs and herbaceous plants whereas Pythium species attacked primarily monocotyledonous herbaceous plants, whereas some cause diseases in fishes, red algae and mammals including humans. However, several mycoparasitic and entomopathogenic species of Pythium have been utilized respectively, to successfully control other plant pathogenic fungi and harmful insects including mosquitoes while the others utilized to produce valuable chemicals for pharmacy and food industry. -

Controlled Sampling of Ribosomally Active Protistan Diversity in Sediment-Surface Layers Identifies Putative Players in the Marine Carbon Sink
The ISME Journal (2020) 14:984–998 https://doi.org/10.1038/s41396-019-0581-y ARTICLE Controlled sampling of ribosomally active protistan diversity in sediment-surface layers identifies putative players in the marine carbon sink 1,2 1 1 3 3 Raquel Rodríguez-Martínez ● Guy Leonard ● David S. Milner ● Sebastian Sudek ● Mike Conway ● 1 1 4,5 6 7 Karen Moore ● Theresa Hudson ● Frédéric Mahé ● Patrick J. Keeling ● Alyson E. Santoro ● 3,8 1,9 Alexandra Z. Worden ● Thomas A. Richards Received: 6 October 2019 / Revised: 4 December 2019 / Accepted: 17 December 2019 / Published online: 9 January 2020 © The Author(s) 2020. This article is published with open access Abstract Marine sediments are one of the largest carbon reservoir on Earth, yet the microbial communities, especially the eukaryotes, that drive these ecosystems are poorly characterised. Here, we report implementation of a sampling system that enables injection of reagents into sediments at depth, allowing for preservation of RNA in situ. Using the RNA templates recovered, we investigate the ‘ribosomally active’ eukaryotic diversity present in sediments close to the water/sediment interface. We 1234567890();,: 1234567890();,: demonstrate that in situ preservation leads to recovery of a significantly altered community profile. Using SSU rRNA amplicon sequencing, we investigated the community structure in these environments, demonstrating a wide diversity and high relative abundance of stramenopiles and alveolates, specifically: Bacillariophyta (diatoms), labyrinthulomycetes and ciliates. The identification of abundant diatom rRNA molecules is consistent with microscopy-based studies, but demonstrates that these algae can also be exported to the sediment as active cells as opposed to dead forms. -

Horizontal Gene Transfer in Osmotrophs: Playing with Public Goods
ORE Open Research Exeter TITLE Horizontal gene transfer in osmotrophs: playing with public goods. AUTHORS Richards, Thomas A; Talbot, Nicholas J. JOURNAL Nat Rev Microbiol DEPOSITED IN ORE 19 November 2014 This version available at http://hdl.handle.net/10871/15898 COPYRIGHT AND REUSE Open Research Exeter makes this work available in accordance with publisher policies. A NOTE ON VERSIONS The version presented here may differ from the published version. If citing, you are advised to consult the published version for pagination, volume/issue and date of publication PERSPECTIVES however, not unique to fungi. Many bacteria, OPINION for instance, feed in an analogous man ner, and other eukaryotic groups, such as Horizontal gene transfer in hyphochytriomycetes (FIG. 1c) and oomycetes (FIG. 1d) (sometimes collectively termed the pseudofungi26), also feed osmotrophically osmotrophs: playing with public and adopt filamentous growth habits, allow ing invasive growth in heterogeneous sub goods strates. Importantly, these eukaryotes also lost the ability to carry out phagotrophy and Thomas A. Richards and Nicholas J. Talbot became obligately osmotrophic26,27. Osmotrophy has a number of distinct Abstract | Osmotrophic microorganisms, such as fungi and oomycetes, feed advantages as a feeding strategy. External by secreting depolymerizing enzymes to process complex food sources in the digestion of large and complex polymers extracellular environment, and taking up the resulting simple sugars, micronutrients allows greater control over substances that and amino acids. As a consequence of this lifestyle, osmotrophs engage in the are allowed to enter a cell (FIG. 2a), thus acquisition and protection of public goods. In this Opinion article, we propose that minimizing potential routes of infection and intake of harmful substances. -

Horizontal Gene Transfer
Horizontal Gene Transfer (HGT) from Fungi is the RESEARCH NEWS basis for plant pathogenicity in oomycetes It always seemed rather odd that some of which seem to have occurred close to that time. This more detailed study, made oomycetes (Oomycota) were so fungal-like the shift from phagotrophy to osmotrophy possible by increasingly available genome in their behaviour as plant pathogens. and the evolution of the fungal cell wall at sequences, reveals that this phenomenon Now whole-genome comparisons are the base of the monophyletic Fungi clade. is much more extensive than might have starting to reveal just why. Richards et al. Amongst the genes evidently transferred, been imagined. Thus, it is not a matter of (2011) have undertaken a painstaking are one related to features such as the ability oomycetes merely being ‘fungal analogues’ gene-by-gene analysis of the proteomes to break down plant cell walls and take or ‘pseudofungi’, they are actually partly of Hyaloperonospora parasitica and three up sugars, nitrogen and phosphates, and Fungi. species of Phytophthora (P. infestans, P. further ones implicated in overcoming plant It should be noted that HGT is not ramorum, and P. sojae) which reveals defence mechanisms and attacking plant only unidirectional from Fungi into to an extensive pattern of cross-kingdom cells. A schematic diagram of the functional other organisms. Indeed, in another paper horizontal gene transfer (HGT) from Fungi. proteome of oomycetes derived from published this summer, this same group of In the case of P. ramorum, an amazing 7.6 Fungi is provided (their Fig. 2) at which researchers report identifying 323 examples % of the secreted proteome appears to have one can only marvel at the complexity. -

Seven Gene Phylogeny of Heterokonts
ARTICLE IN PRESS Protist, Vol. 160, 191—204, May 2009 http://www.elsevier.de/protis Published online date 9 February 2009 ORIGINAL PAPER Seven Gene Phylogeny of Heterokonts Ingvild Riisberga,d,1, Russell J.S. Orrb,d,1, Ragnhild Klugeb,c,2, Kamran Shalchian-Tabrizid, Holly A. Bowerse, Vishwanath Patilb,c, Bente Edvardsena,d, and Kjetill S. Jakobsenb,d,3 aMarine Biology, Department of Biology, University of Oslo, P.O. Box 1066, Blindern, NO-0316 Oslo, Norway bCentre for Ecological and Evolutionary Synthesis (CEES),Department of Biology, University of Oslo, P.O. Box 1066, Blindern, NO-0316 Oslo, Norway cDepartment of Plant and Environmental Sciences, P.O. Box 5003, The Norwegian University of Life Sciences, N-1432, A˚ s, Norway dMicrobial Evolution Research Group (MERG), Department of Biology, University of Oslo, P.O. Box 1066, Blindern, NO-0316, Oslo, Norway eCenter of Marine Biotechnology, 701 East Pratt Street, Baltimore, MD 21202, USA Submitted May 23, 2008; Accepted November 15, 2008 Monitoring Editor: Mitchell L. Sogin Nucleotide ssu and lsu rDNA sequences of all major lineages of autotrophic (Ochrophyta) and heterotrophic (Bigyra and Pseudofungi) heterokonts were combined with amino acid sequences from four protein-coding genes (actin, b-tubulin, cox1 and hsp90) in a multigene approach for resolving the relationship between heterokont lineages. Applying these multigene data in Bayesian and maximum likelihood analyses improved the heterokont tree compared to previous rDNA analyses by placing all plastid-lacking heterotrophic heterokonts sister to Ochrophyta with robust support, and divided the heterotrophic heterokonts into the previously recognized phyla, Bigyra and Pseudofungi. Our trees identified the heterotrophic heterokonts Bicosoecida, Blastocystis and Labyrinthulida (Bigyra) as the earliest diverging lineages. -

Kingdom Chromista)
J Mol Evol (2006) 62:388–420 DOI: 10.1007/s00239-004-0353-8 Phylogeny and Megasystematics of Phagotrophic Heterokonts (Kingdom Chromista) Thomas Cavalier-Smith, Ema E-Y. Chao Department of Zoology, University of Oxford, South Parks Road, Oxford OX1 3PS, UK Received: 11 December 2004 / Accepted: 21 September 2005 [Reviewing Editor: Patrick J. Keeling] Abstract. Heterokonts are evolutionarily important gyristea cl. nov. of Ochrophyta as once thought. The as the most nutritionally diverse eukaryote supergroup zooflagellate class Bicoecea (perhaps the ancestral and the most species-rich branch of the eukaryotic phenotype of Bigyra) is unexpectedly diverse and a kingdom Chromista. Ancestrally photosynthetic/ major focus of our study. We describe four new bicil- phagotrophic algae (mixotrophs), they include several iate bicoecean genera and five new species: Nerada ecologically important purely heterotrophic lineages, mexicana, Labromonas fenchelii (=Pseudobodo all grossly understudied phylogenetically and of tremulans sensu Fenchel), Boroka karpovii (=P. uncertain relationships. We sequenced 18S rRNA tremulans sensu Karpov), Anoeca atlantica and Cafe- genes from 14 phagotrophic non-photosynthetic het- teria mylnikovii; several cultures were previously mis- erokonts and a probable Ochromonas, performed ph- identified as Pseudobodo tremulans. Nerada and the ylogenetic analysis of 210–430 Heterokonta, and uniciliate Paramonas are related to Siluania and revised higher classification of Heterokonta and its Adriamonas; this clade (Pseudodendromonadales three phyla: the predominantly photosynthetic Och- emend.) is probably sister to Bicosoeca. Genetically rophyta; the non-photosynthetic Pseudofungi; and diverse Caecitellus is probably related to Anoeca, Bigyra (now comprising subphyla Opalozoa, Bicoecia, Symbiomonas and Cafeteria (collectively Anoecales Sagenista). The deepest heterokont divergence is emend.). Boroka is sister to Pseudodendromonadales/ apparently between Bigyra, as revised here, and Och- Bicoecales/Anoecales. -
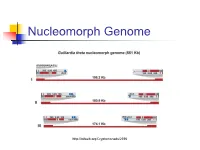
Nucleomorph Genome
Nucleomorph Genome http://tolweb.org/Cryptomonads/2396 Crypt of the cryptophytes V – Vestibulum (vestíbulo) F – Furrow (sulco) G – Gullet (garganta) Brett et al. (1994) Protoplasma 181, 106. Types of periplasts Brett et al. (1994) Protoplasma 181, 106. Features of the cryptophytes • Biflagellated cells • Live in marine and freshwater habitats • Survive at low light levels • They have phycobiliproteins • Great importance in the food chain • They have chloroplasts with 4 membranes • They have a crypt that is structurally supported by a periplast • They have chlorophyll c2 instead of chlorophyll b • They do not have allophycocyanin • Periplastidial starch • Thylakoids grouped in pairs Cryptophytes vs. Chloraracniophytes Chlorarachniophyta Cryptophyta Photosynthetic pigments: Photosynthetic pigments: chlorophyll a and chlorophyll b chlorophyll a and c2 as well as other pigments evolutionarily related to phycoerythrins Do not have phycobilissomes Biliproteins within the thylakoid or biliproteins lumen; thylakoids grouped in pairs Uniflagellar ameboid cells with 2 flagella inserted near a crypt filamentous pseudopodia and asymmetric ellipsoidal cells Starch in cytoplasmic vesicles Periplastic starch (between the plastid envelope and the two outer membranes) Chloroplast surrounded by 4 Chloroplast surrounded by 4 membranes membranes ER not associated with the ER associated with the chloroplast chloroplast http://www.laborjournal.de/blog/?p=5529 Lineage Stramenopiles http://tolweb.org/Eukaryotes/3 Lineage Heterokonts Lineage Heterokonts • Heterokonts ≈ Stramenopiles • Biflagellated eukaryotic cells • Previous flagellum with mastigonemes (tripartite tubular hairs) • Hairless posterior flagellum • Chloroplasts surrounded by 4 membranes (photoautotrophic sub-lineage: Ochrophyta ≈ Heterokontophyta) • However, there are cells that have lost one or more flagella Lineage Heterokonts • Diatoms (> 100,000 species) • Brown seaweeds (Phaeophyceae) • Chrysophytes (golden algae) • Pseudofungi: Oomycetes (heterotrophic sub-lineage) • etc.. -

Horizontal Gene Transfer Facilitated the Evolution of Plant Parasitic Mechanisms in the Oomycetes
Horizontal gene transfer facilitated the evolution of plant parasitic mechanisms in the oomycetes Thomas A. Richardsa,b,1, Darren M. Soanesa, Meredith D. M. Jonesa,b, Olga Vasievac, Guy Leonarda,b, Konrad Paszkiewicza, Peter G. Fosterb, Neil Hallc, and Nicholas J. Talbota aBiosciences, University of Exeter, Exeter EX4 4QD, United Kingdom; bDepartment of Zoology, Natural History Museum, London SW7 5BD, United Kingdom; and cSchool of Biological Sciences, University of Liverpool, Liverpool L69 7ZB, United Kingdom Edited by W. Ford Doolittle, Dalhousie University, Halifax, Canada, and approved July 27, 2011 (received for review March 31, 2011) Horizontal gene transfer (HGT) can radically alter the genomes of ramorum, for example, whereas the Irish potato famine of the microorganisms, providing the capacity to adapt to new lifestyles, 19th century was caused by the late blight parasite Phytophthora environments, and hosts. However, the extent of HGT between infestans. Important crop diseases caused by fungi include the eukaryotes is unclear. Using whole-genome, gene-by-gene phylo- devastating rice blast disease caused by M. oryzae and the rusts, genetic analysis we demonstrate an extensive pattern of cross- smuts, and mildews that affect wheat, barley, and maize. In this kingdom HGT between fungi and oomycetes. Comparative study we report that HGT between fungi and oomycetes has genomics, including the de novo genome sequence of Hyphochy- occurred to a far greater degree than hitherto recognized (19). trium catenoides, a free-living sister of the oomycetes, shows that Our previous analysis suggested four strongly supported cases of these transfers largely converge within the radiation of oomycetes HGT, but by using a whole-genome, gene-by-gene phylogenetic that colonize plant tissues. -

Genome Assembly of Nannochloropsis Oceanica Provides Evidence of Host Nucleus Overthrow by the Symbiont Nucleus During Speciation
ARTICLE https://doi.org/10.1038/s42003-019-0500-9 OPEN Genome assembly of Nannochloropsis oceanica provides evidence of host nucleus overthrow by the symbiont nucleus during speciation Li Guo1, Sijie Liang1, Zhongyi Zhang1, Hang Liu1, Songwen Wang2, Kehou Pan3, Jian Xu4, Xue Ren5, Surui Pei5 & 1234567890():,; Guanpin Yang1,6,7 The species of the genus Nannochloropsis are unique in their maintenance of a nucleus-plastid continuum throughout their cell cycle, non-motility and asexual reproduction. These char- acteristics should have been endorsed in their gene assemblages (genomes). Here we show that N. oceanica has a genome of 29.3 Mb consisting of 32 pseudochromosomes and con- taining 7,330 protein-coding genes; and the host nucleus may have been overthrown by an ancient red alga symbiont nucleus during speciation through secondary endosymbiosis. In addition, N. oceanica has lost its flagella and abilities to undergo meiosis and sexual repro- duction, and adopted a genome reduction strategy during speciation. We propose that N. oceanica emerged through the active fusion of a host protist and a photosynthesizing ancient red alga and the symbiont nucleus became dominant over the host nucleus while the chloroplast was wrapped by two layers of endoplasmic reticulum. Our findings evidenced an alternative speciation pathway of eukaryotes. 1 College of Marine Life Sciences, Ocean University of China (OUC), Qingdao 266003, P. R. China. 2 College of Agriculture and Resources and Environment, Tianjin Agricultural University, Tianjin 300384, P. R. China. 3 Laboratory of Applied Microalgae, College of Fisheries, OUC, Qingdao 266003, P. R. China. 4 Functional Genomics Group, Qingdao Institute of Bioenergy and Bioprocess Technology, Chinese Academy of Sciences, Qingdao 266101, P. -

Genera of the World: an Overview and Estimates Based on the March 2020 Release of the Interim Register of Marine and Nonmarine Genera (IRMNG)
Megataxa 001 (2): 123–140 ISSN 2703-3082 (print edition) https://www.mapress.com/j/mt/ MEGATAXA Copyright © 2020 Magnolia Press Article ISSN 2703-3090 (online edition) https://doi.org/10.11646/megataxa.1.2.3 http://zoobank.org/urn:lsid:zoobank.org:pub:F4A52C97-BAD0-4FD5-839F-1A61EA40A7A3 All genera of the world: an overview and estimates based on the March 2020 release of the Interim Register of Marine and Nonmarine Genera (IRMNG) TONY REES 1, LEEN VANDEPITTE 2, 3, BART VANHOORNE 2, 4 & WIM DECOCK 2, 5 1 Private address, New South Wales, Australia. �[email protected]; http://orcid.org/0000-0003-1887-5211 2 Flanders Marine Institute/Vlaams Instituut Voor De Zee (VLIZ), Wandelaarkaai 7, 8400 Ostend, Belgium. 3 �[email protected]; http://orcid.org/0000-0002-8160-7941 4 �[email protected]; https://orcid.org/0000-0002-6642-4725 5 �[email protected]; https://orcid.org/0000-0002-2168-9471 Abstract Introduction We give estimated counts of known accepted genera of the The concept of a series of papers addressing portions of world (297,930±65,840, of which approximately 21% are the question of “all genera of the world” is a valuable one, fossil), of a total 492,620 genus names presently held for which can benefit from as much preliminary scoping as “all life”, based on the March 2020 release of the Interim may be currently available. To date, synoptic surveys of Register of Marine and Nonmarine Genera (IRMNG). A fur- biodiversity have been attempted mainly at the level of ther c. -

The Chromista
Maneveldt, G.W. & Keats, D.W. (1997). The chromista. ENGINEERING IN LIFE SCIENCES The Chromista Maneveldt, Gavin W. Department of Biodiversity and Conservation Biology, University of the Western Cape South Africa Keats, Derek W. Department of Information and Communication Services, University of the Western Cape South Africa As a group, the chromists show a diverse range of forms from tiny unicellular, flagellates to the large brown algae known as kelp. Molecular studies have confirmed the inclusion of certain organisms once considered Fungi, as well as some heterotrophic flagellates. Despite their diversity of form and feeding modes, a few unique characters group these organisms. Introduction The 5-Kingdom system of Robert H. Whittaker (see Whittaker, 1959; 1969), although a major improvement over the simple 2-Kingdom system of plants and animals, still left much room for criticism (see South and Whittick, 1987 for a review). While the Kingdom Monera, with its distinct prokaryote organization of the nucleus, is still considered the most natural and unambiguous Kingdom of this system, the remaining Kingdoms have all undergone some or other taxonomic scrutiny. The Plant, Animal and Fungi Kingdoms, all multicellular eukaryotes, have each been defined by their nutritional modes. Several researchers have criticized the use of these characters arguing that they defined adaptive traits and not taxa. These names are thus entirely unsuitable for revealing phylogenetic relationships among organisms. The Kingdom Protista or Protoctista is the hardest to define, as it is the least homogenous group. It is somewhat of a mix-n-match containing all eukaryotes that do not fit into the plant, animal or fungi kingdoms.