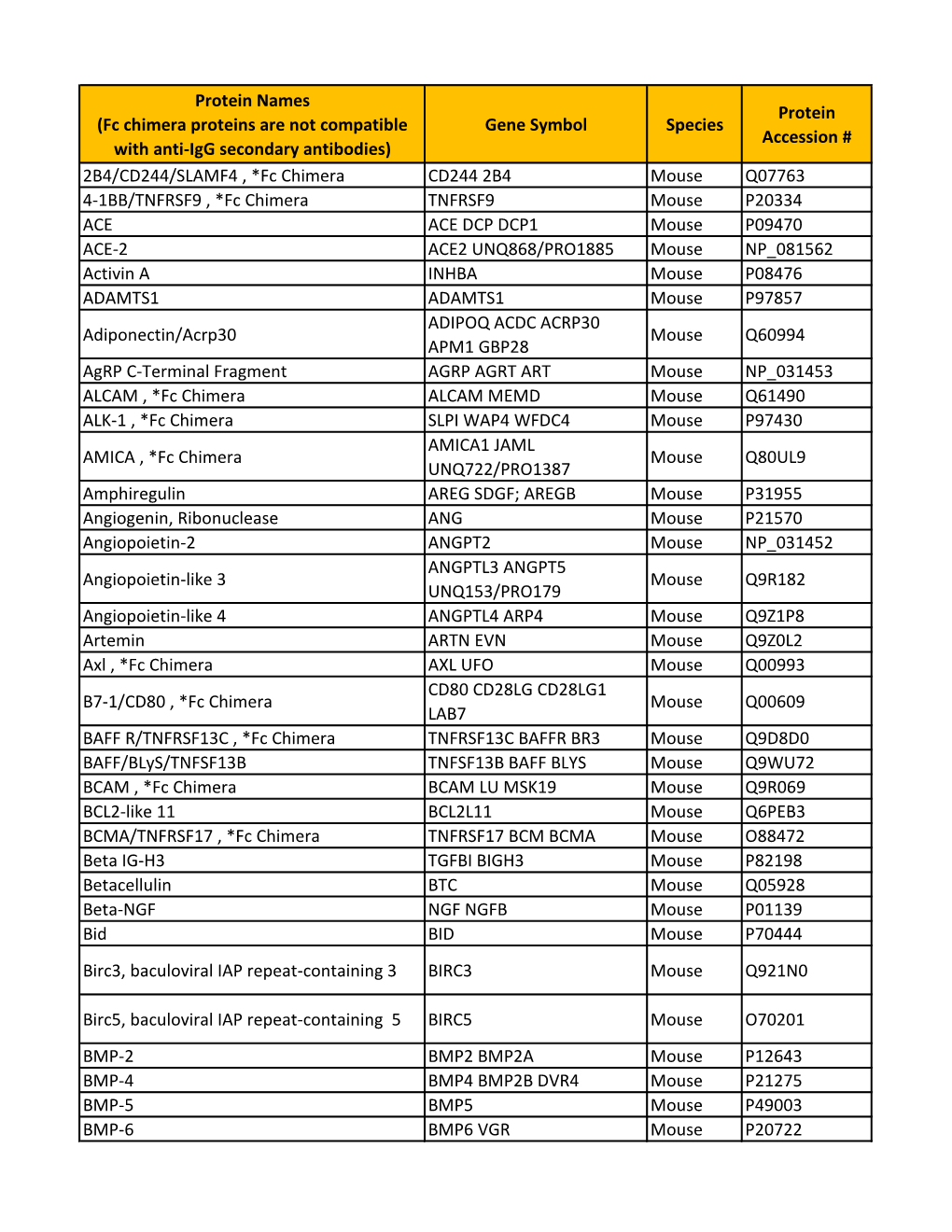
Gene Symbol Species Protein Accession
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Increased Expression of Epidermal Growth Factor Receptor and Betacellulin During the Early Stage of Gastric Ulcer Healing
505-510 11/6/08 12:55 Page 505 MOLECULAR MEDICINE REPORTS 1: 505-510, 2008 505 Increased expression of epidermal growth factor receptor and betacellulin during the early stage of gastric ulcer healing GEUN HAE CHOI, HO SUNG PARK, KYUNG RYOUL KIM, HA NA CHOI, KYU YUN JANG, MYOUNG JA CHUNG, MYOUNG JAE KANG, DONG GEUN LEE and WOO SUNG MOON Department of Pathology, Institute for Medical Sciences, Chonbuk National University Medical School and the Center for Healthcare Technology Development, Jeonju, Korea Received January 2, 2008; Accepted February 22, 2008 Abstract. Epidermal growth factor receptor (EGFR) is from tissue necrosis triggered by mucosal ischemia, free important for the proliferation and differentiation of gastric radical formation and the cessation of nutrient delivery, which mucosal cells. Betacellulin (BTC) is a novel ligand for EGFR are caused by vascular and microvascular injury such as Since their role is unclear in the ulcer healing process, we thrombi, constriction or other occlusions (2). Tissue necrosis investigated their expression. Gastric ulcers in 30 Sprague- and the release of leukotriene B attract leukocytes and Dawley rats were induced by acetic acid. RT-PCR and macrophages, which release pro-inflammatory cytokines Western blotting were performed to detect EGFR and BTC. (e.g. TNFα, IL-1α, and IL-1ß). These in turn activate local Immunohistochemical studies were performed to detect fibroblasts, endothelial and epithelial cells. Histologically, an EGFR, BTC and proliferating cell nuclear antigen (PCNA). ulcer has two characteristic structures: a distinct ulcer margin The expression of EGFR and the BTC gene was significantly formed by the adjacent non-necrotic mucosa, and granulation increased at 12 h, 24 h and 3 days after ulcer induction tissue composed of fibroblasts, macrophages and proliferating (P<0.05). -

The TNF and TNF Receptor Review Superfamilies: Integrating Mammalian Biology
Cell, Vol. 104, 487±501, February 23, 2001, Copyright 2001 by Cell Press The TNF and TNF Receptor Review Superfamilies: Integrating Mammalian Biology Richard M. Locksley,*²³k Nigel Killeen,²k The receptors and ligands in this superfamily have and Michael J. Lenardo§k unique structural attributes that couple them directly to *Department of Medicine signaling pathways for cell proliferation, survival, and ² Department of Microbiology and Immunology differentiation. Thus, they have assumed prominent ³ Howard Hughes Medical Institute roles in the generation of tissues and transient microen- University of California, San Francisco vironments. Most TNF/TNFR SFPs are expressed in the San Francisco, California 94143 immune system, where their rapid and potent signaling § Laboratory of Immunology capabilities are crucial in coordinating the proliferation National Institute of Allergy and Infectious Diseases and protective functions of pathogen-reactive cells. National Institutes of Health Here, we review the organization of the TNF/TNFR SF Bethesda, Maryland 20892 and how these proteins have been adapted for pro- cesses as seemingly disparate as host defense and or- ganogenesis. In interpreting this large and highly active Introduction area of research, we have focused on common themes that unite the actions of these genes in different tissues. Three decades ago, lymphotoxin (LT) and tumor necro- We also discuss the evolutionary success of this super- sis factor (TNF) were identified as products of lympho- familyÐsuccess that we infer from its expansion across cytes and macrophages that caused the lysis of certain the mammalian genome and from its many indispens- types of cells, especially tumor cells (Granger et al., able roles in mammalian biology. -

Fog2 Is Critical for Cardiac Function and Maintenance of Coronary Vasculature in the Adult Mouse Heart
Fog2 is critical for cardiac function and maintenance of coronary vasculature in the adult mouse heart Bin Zhou, … , Sergei G. Tevosian, William T. Pu J Clin Invest. 2009;119(6):1462-1476. https://doi.org/10.1172/JCI38723. Research Article Cardiology Aberrant transcriptional regulation contributes to the pathogenesis of both congenital and adult forms of heart disease. While the transcriptional regulator friend of Gata 2 (FOG2) is known to be essential for heart morphogenesis and coronary development, its tissue-specific function has not been previously investigated. Additionally, little is known about the role of FOG2 in the adult heart. Here we used spatiotemporally regulated inactivation of Fog2 to delineate its function in both the embryonic and adult mouse heart. Early cardiomyocyte-restricted loss of Fog2 recapitulated the cardiac and coronary defects of the Fog2 germline murine knockouts. Later cardiomyocyte-restricted loss ofF og2 (Fog2MC) did not result in defects in cardiac structure or coronary vessel formation. However, Fog2MC adult mice had severely depressed ventricular function and died at 8–14 weeks. Fog2MC adult hearts displayed a paucity of coronary vessels, associated with myocardial hypoxia, increased cardiomyocyte apoptosis, and cardiac fibrosis. Induced inactivation of Fog2 in the adult mouse heart resulted in similar phenotypes, as did ablation of the FOG2 interaction with the transcription factor GATA4. Loss of the FOG2 or FOG2-GATA4 interaction altered the expression of a panel of angiogenesis-related genes. Collectively, our data indicate that FOG2 regulates adult heart function and coronary angiogenesis. Find the latest version: https://jci.me/38723/pdf Research article Fog2 is critical for cardiac function and maintenance of coronary vasculature in the adult mouse heart Bin Zhou,1,2 Qing Ma,1 Sek Won Kong,1 Yongwu Hu,1,3 Patrick H. -

Recombinant Human Betacellulin Promotes the Neogenesis of -Cells
Recombinant Human Betacellulin Promotes the Neogenesis of -Cells and Ameliorates Glucose Intolerance in Mice With Diabetes Induced by Selective Alloxan Perfusion Koji Yamamoto, Jun-ichiro Miyagawa, Masako Waguri, Reiko Sasada, Koichi Igarashi, Ming Li, Takao Nammo, Makoto Moriwaki, Akihisa Imagawa, Kazuya Yamagata, Hiromu Nakajima, Mitsuyoshi Namba, Yoshihiro Tochino, Toshiaki Hanafusa, and Yuji Matsuzawa Betacellulin (BTC), a member of the epidermal growth factor family, is expressed predominantly in the human pancreas and induces the differentiation of a pancreatic ancreatic -cells are thought to be terminally dif- acinar cell line (AR42J) into insulin-secreting cells, ferentiated cells with little ability to regenerate. suggesting that BTC has a physiologically important However, proliferation of preexisting -cells and role in the endocrine pancreas. In this study, we exam- differentiation of -cells from precursor cells, ined the in vivo effect of recombinant human BTC P (rhBTC) on glucose intolerance and pancreatic mor- mainly residing in the pancreatic duct lining, have been phology using a new mouse model with glucose intoler- demonstrated in some animal models (1–5). Recently, we ance induced by selective alloxan perfusion. RhBTC developed a new mouse model of diabetes induced by selec- (1 µg/g body wt) or saline was injected subcutaneously tive perfusion of alloxan (100 µg/g body wt) during the every day from the day after alloxan treatment. The clamping of the superior mesenteric artery (1). In this model, intraperitoneal glucose tolerance test revealed no dif- glucose intolerance spontaneously resolves after one year ference between rhBTC-treated and rhBTC-untreated because of the proliferation of surviving -cells in the non- glucose-intolerant mice at 2–4 weeks. -

Tools for Cell Therapy and Immunoregulation
RnDSy-lu-2945 Tools for Cell Therapy and Immunoregulation Target Cell TIM-4 SLAM/CD150 BTNL8 PD-L2/B7-DC B7-H1/PD-L1 (Human) Unknown PD-1 B7-1/CD80 TIM-1 SLAM/CD150 Receptor TIM Family SLAM Family Butyrophilins B7/CD28 Families T Cell Multiple Co-Signaling Molecules Co-stimulatory Co-inhibitory Ig Superfamily Regulate T Cell Activation Target Cell T Cell Target Cell T Cell B7-1/CD80 B7-H1/PD-L1 T cell activation requires two signals: 1) recognition of the antigenic peptide/ B7-1/CD80 B7-2/CD86 CTLA-4 major histocompatibility complex (MHC) by the T cell receptor (TCR) and 2) CD28 antigen-independent co-stimulation induced by interactions between B7-2/CD86 B7-H1/PD-L1 B7-1/CD80 co-signaling molecules expressed on target cells, such as antigen-presenting PD-L2/B7-DC PD-1 ICOS cells (APCs), and their T cell-expressed receptors. Engagement of the TCR in B7-H2/ICOS L 2Ig B7-H3 (Mouse) the absence of this second co-stimulatory signal typically results in T cell B7-H1/PD-L1 B7/CD28 Families 4Ig B7-H3 (Human) anergy or apoptosis. In addition, T cell activation can be negatively regulated Unknown Receptors by co-inhibitory molecules present on APCs. Therefore, integration of the 2Ig B7-H3 Unknown B7-H4 (Mouse) Receptors signals transduced by co-stimulatory and co-inhibitory molecules following TCR B7-H5 4Ig B7-H3 engagement directs the outcome and magnitude of a T cell response Unknown Ligand (Human) B7-H5 including the enhancement or suppression of T cell proliferation, B7-H7 Unknown Receptor differentiation, and/or cytokine secretion. -

Cytokine Nomenclature
RayBiotech, Inc. The protein array pioneer company Cytokine Nomenclature Cytokine Name Official Full Name Genbank Related Names Symbol 4-1BB TNFRSF Tumor necrosis factor NP_001552 CD137, ILA, 4-1BB ligand receptor 9 receptor superfamily .2. member 9 6Ckine CCL21 6-Cysteine Chemokine NM_002989 Small-inducible cytokine A21, Beta chemokine exodus-2, Secondary lymphoid-tissue chemokine, SLC, SCYA21 ACE ACE Angiotensin-converting NP_000780 CD143, DCP, DCP1 enzyme .1. NP_690043 .1. ACE-2 ACE2 Angiotensin-converting NP_068576 ACE-related carboxypeptidase, enzyme 2 .1 Angiotensin-converting enzyme homolog ACTH ACTH Adrenocorticotropic NP_000930 POMC, Pro-opiomelanocortin, hormone .1. Corticotropin-lipotropin, NPP, NP_001030 Melanotropin gamma, Gamma- 333.1 MSH, Potential peptide, Corticotropin, Melanotropin alpha, Alpha-MSH, Corticotropin-like intermediary peptide, CLIP, Lipotropin beta, Beta-LPH, Lipotropin gamma, Gamma-LPH, Melanotropin beta, Beta-MSH, Beta-endorphin, Met-enkephalin ACTHR ACTHR Adrenocorticotropic NP_000520 Melanocortin receptor 2, MC2-R hormone receptor .1 Activin A INHBA Activin A NM_002192 Activin beta-A chain, Erythroid differentiation protein, EDF, INHBA Activin B INHBB Activin B NM_002193 Inhibin beta B chain, Activin beta-B chain Activin C INHBC Activin C NM005538 Inhibin, beta C Activin RIA ACVR1 Activin receptor type-1 NM_001105 Activin receptor type I, ACTR-I, Serine/threonine-protein kinase receptor R1, SKR1, Activin receptor-like kinase 2, ALK-2, TGF-B superfamily receptor type I, TSR-I, ACVRLK2 Activin RIB ACVR1B -

Serum Protein Fingerprinting by PEA Immunoassay Coupled with a Pattern-Recognition Algorithms Distinguishes MGUS and Multiple Myeloma
www.impactjournals.com/oncotarget/ Oncotarget, 2017, Vol. 8, (No. 41), pp: 69408-69421 Research Paper Serum protein fingerprinting by PEA immunoassay coupled with a pattern-recognition algorithms distinguishes MGUS and multiple myeloma Petra Schneiderova1,*, Tomas Pika2,*, Petr Gajdos3, Regina Fillerova1, Pavel Kromer3, Milos Kudelka3, Jiri Minarik2, Tomas Papajik2, Vlastimil Scudla2 and Eva Kriegova1 1Department of Immunology, Faculty of Medicine and Dentistry, Palacky University Olomouc, Olomouc, Czech Republic 2Department of Hemato-Oncology, Faculty of Medicine and Dentistry, Palacky University and University Hospital, Olomouc, Czech Republic 3Department of Computer Science, Faculty of Electrical Engineering and Computer Science, VSB-Technical University of Ostrava, Ostrava, Czech Republic *These authors have contributed equally to this work Correspondence to: Eva Kriegova, email: [email protected] Keywords: serum pattern, cytokines, growth factors, proximity extension immunoassay, post-transplant serum pattern Received: May 04, 2016 Accepted: July 28, 2016 Published: August 12, 2016 Copyright: Schneiderova et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License 3.0 (CC BY 3.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited. ABSTRACT Serum protein fingerprints associated with MGUS and MM and their changes in MM after autologous stem cell transplantation (MM-ASCT, day 100) remain unexplored. Using highly-sensitive Proximity Extension ImmunoAssay on 92 cancer biomarkers (Proseek Multiplex, Olink), enhanced serum levels of Adrenomedullin (ADM, Pcorr= .0004), Growth differentiation factor 15 (GDF15, Pcorr= .003), and soluble Major histocompatibility complex class I-related chain A (sMICA, Pcorr= .023), all prosurvival and chemoprotective factors for myeloma cells, were detected in MM comparing to MGUS. -

Supplemental Materials ZNF281 Enhances Cardiac Reprogramming
Supplemental Materials ZNF281 enhances cardiac reprogramming by modulating cardiac and inflammatory gene expression Huanyu Zhou, Maria Gabriela Morales, Hisayuki Hashimoto, Matthew E. Dickson, Kunhua Song, Wenduo Ye, Min S. Kim, Hanspeter Niederstrasser, Zhaoning Wang, Beibei Chen, Bruce A. Posner, Rhonda Bassel-Duby and Eric N. Olson Supplemental Table 1; related to Figure 1. Supplemental Table 2; related to Figure 1. Supplemental Table 3; related to the “quantitative mRNA measurement” in Materials and Methods section. Supplemental Table 4; related to the “ChIP-seq, gene ontology and pathway analysis” and “RNA-seq” and gene ontology analysis” in Materials and Methods section. Supplemental Figure S1; related to Figure 1. Supplemental Figure S2; related to Figure 2. Supplemental Figure S3; related to Figure 3. Supplemental Figure S4; related to Figure 4. Supplemental Figure S5; related to Figure 6. Supplemental Table S1. Genes included in human retroviral ORF cDNA library. Gene Gene Gene Gene Gene Gene Gene Gene Symbol Symbol Symbol Symbol Symbol Symbol Symbol Symbol AATF BMP8A CEBPE CTNNB1 ESR2 GDF3 HOXA5 IL17D ADIPOQ BRPF1 CEBPG CUX1 ESRRA GDF6 HOXA6 IL17F ADNP BRPF3 CERS1 CX3CL1 ETS1 GIN1 HOXA7 IL18 AEBP1 BUD31 CERS2 CXCL10 ETS2 GLIS3 HOXB1 IL19 AFF4 C17ORF77 CERS4 CXCL11 ETV3 GMEB1 HOXB13 IL1A AHR C1QTNF4 CFL2 CXCL12 ETV7 GPBP1 HOXB5 IL1B AIMP1 C21ORF66 CHIA CXCL13 FAM3B GPER HOXB6 IL1F3 ALS2CR8 CBFA2T2 CIR1 CXCL14 FAM3D GPI HOXB7 IL1F5 ALX1 CBFA2T3 CITED1 CXCL16 FASLG GREM1 HOXB9 IL1F6 ARGFX CBFB CITED2 CXCL3 FBLN1 GREM2 HOXC4 IL1F7 -

Human B-Cell Proliferation and Intracellular Signaling: Part 3
1872 Diabetes Volume 64, June 2015 Andrew F. Stewart,1 Mehboob A. Hussain,2 Adolfo García-Ocaña,1 Rupangi C. Vasavada,1 Anil Bhushan,3 Ernesto Bernal-Mizrachi,4 and Rohit N. Kulkarni5 Human b-Cell Proliferation and Intracellular Signaling: Part 3 Diabetes 2015;64:1872–1885 | DOI: 10.2337/db14-1843 This is the third in a series of Perspectives on intracel- signaling pathways in rodent and human b-cells, with lular signaling pathways coupled to proliferation in pan- a specific focus on the links between b-cell proliferation creatic b-cells. We contrast the large knowledge base in and intracellular signaling pathways (1,2). We highlight rodent b-cells with the more limited human database. what is known in rodent b-cells and compare and contrast With the increasing incidence of type 1 diabetes and that to the current knowledge base in human b-cells. In- the recognition that type 2 diabetes is also due in part variably, the human b-cell section is very brief compared fi b to a de ciency of functioning -cells, there is great ur- with the rodent counterpart, reflecting the still primitive gency to identify therapeutic approaches to expand hu- state of our understanding of mitogenic signaling in hu- b man -cell numbers. Therapeutic approaches might man b-cells. To emphasize this difference, each figure is include stem cell differentiation, transdifferentiation, or divided into two panels, one summarizing rodent b-cell expansion of cadaver islets or residual endogenous signaling and one for human b-cells. Our intended audi- b-cells. In these Perspectives, we focus on b-cell ence includes trainees in b-cell regeneration as well as proliferation. -

Supplementary Material
Supplementary Material Table S1: Significant downregulated KEGGs pathways identified by DAVID following exposure to five cinnamon- based phenylpropanoids (p < 0.05). p-value Term: Genes (Benjamini) Cytokine-cytokine receptor interaction: FASLG, TNFSF14, CXCL11, IL11, FLT3LG, CCL3L1, CCL3L3, CXCR6, XCR1, 2.43 × 105 RTEL1, CSF2RA, TNFRSF17, TNFRSF14, CCNL2, VEGFB, AMH, TNFRSF10B, INHBE, IFNB1, CCR3, VEGFA, CCR2, IL12A, CCL1, CCL3, CXCL5, TNFRSF25, CCR1, CSF1, CX3CL1, CCL7, CCL24, TNFRSF1B, IL12RB1, CCL21, FIGF, EPO, IL4, IL18R1, FLT1, TGFBR1, EDA2R, HGF, TNFSF8, KDR, LEP, GH2, CCL13, EPOR, XCL1, IFNA16, XCL2 Neuroactive ligand-receptor interaction: OPRM1, THRA, GRIK1, DRD2, GRIK2, TACR2, TACR1, GABRB1, LPAR4, 9.68 × 105 GRIK5, FPR1, PRSS1, GNRHR, FPR2, EDNRA, AGTR2, LTB4R, PRSS2, CNR1, S1PR4, CALCRL, TAAR5, GABRE, PTGER1, GABRG3, C5AR1, PTGER3, PTGER4, GABRA6, GABRA5, GRM1, PLG, LEP, CRHR1, GH2, GRM3, SSTR2, Chlorogenic acid Chlorogenic CHRM3, GRIA1, MC2R, P2RX2, TBXA2R, GHSR, HTR2C, TSHR, LHB, GLP1R, OPRD1 Hematopoietic cell lineage: IL4, CR1, CD8B, CSF1, FCER2, GYPA, ITGA2, IL11, GP9, FLT3LG, CD38, CD19, DNTT, 9.29 × 104 GP1BB, CD22, EPOR, CSF2RA, CD14, THPO, EPO, HLA-DRA, ITGA2B Cytokine-cytokine receptor interaction: IL6ST, IL21R, IL19, TNFSF15, CXCR3, IL15, CXCL11, TGFB1, IL11, FLT3LG, CXCL10, CCR10, XCR1, RTEL1, CSF2RA, IL21, CCNL2, VEGFB, CCR8, AMH, TNFRSF10C, IFNB1, PDGFRA, EDA, CXCL5, TNFRSF25, CSF1, IFNW1, CNTFR, CX3CL1, CCL5, TNFRSF4, CCL4, CCL27, CCL24, CCL25, CCL23, IFNA6, IFNA5, FIGF, EPO, AMHR2, IL2RA, FLT4, TGFBR2, EDA2R, -

Molecular Mediators of Acute and Chronic Itch in Mouse and Human Sensory Neurons Manouela Valtcheva Washington University in St
Washington University in St. Louis Washington University Open Scholarship Arts & Sciences Electronic Theses and Dissertations Arts & Sciences Spring 5-15-2018 Molecular Mediators of Acute and Chronic Itch in Mouse and Human Sensory Neurons Manouela Valtcheva Washington University in St. Louis Follow this and additional works at: https://openscholarship.wustl.edu/art_sci_etds Part of the Neuroscience and Neurobiology Commons Recommended Citation Valtcheva, Manouela, "Molecular Mediators of Acute and Chronic Itch in Mouse and Human Sensory Neurons" (2018). Arts & Sciences Electronic Theses and Dissertations. 1596. https://openscholarship.wustl.edu/art_sci_etds/1596 This Dissertation is brought to you for free and open access by the Arts & Sciences at Washington University Open Scholarship. It has been accepted for inclusion in Arts & Sciences Electronic Theses and Dissertations by an authorized administrator of Washington University Open Scholarship. For more information, please contact [email protected]. WASHINGTON UNIVERSITY IN ST. LOUIS Division of Biology and Biomedical Sciences Neurosciences Dissertation Examination Committee: Robert W. Gereau, IV, Chair Yu-Qing Cao Sanjay Jain Qin Liu Durga Mohapatra Molecular Mediators of Acute and Chronic Itch in Mouse and Human Sensory Neurons by Manouela Vesselinova Valtcheva A dissertation presented to The Graduate School of Washington University in partial fulfillment of the requirements for the degree of Doctor of Philosophy May 2018 St. Louis, Missouri © 2018, Manouela V. Valtcheva Table -

Angiocrine Endothelium: from Physiology to Cancer Jennifer Pasquier1,2*, Pegah Ghiabi2, Lotf Chouchane3,4,5, Kais Razzouk1, Shahin Rafi3 and Arash Rafi1,2,3
Pasquier et al. J Transl Med (2020) 18:52 https://doi.org/10.1186/s12967-020-02244-9 Journal of Translational Medicine REVIEW Open Access Angiocrine endothelium: from physiology to cancer Jennifer Pasquier1,2*, Pegah Ghiabi2, Lotf Chouchane3,4,5, Kais Razzouk1, Shahin Rafi3 and Arash Rafi1,2,3 Abstract The concept of cancer as a cell-autonomous disease has been challenged by the wealth of knowledge gathered in the past decades on the importance of tumor microenvironment (TM) in cancer progression and metastasis. The sig- nifcance of endothelial cells (ECs) in this scenario was initially attributed to their role in vasculogenesis and angiogen- esis that is critical for tumor initiation and growth. Nevertheless, the identifcation of endothelial-derived angiocrine factors illustrated an alternative non-angiogenic function of ECs contributing to both physiological and pathological tissue development. Gene expression profling studies have demonstrated distinctive expression patterns in tumor- associated endothelial cells that imply a bilateral crosstalk between tumor and its endothelium. Recently, some of the molecular determinants of this reciprocal interaction have been identifed which are considered as potential targets for developing novel anti-angiocrine therapeutic strategies. Keywords: Angiocrine, Endothelium, Cancer, Cancer microenvironment, Angiogenesis Introduction of blood vessels in initiation of tumor growth and stated Metastatic disease accounts for about 90% of patient that in the absence of such angiogenesis, tumors can- mortality. Te difculty in controlling and eradicating not expand their mass or display a metastatic phenotype metastasis might be related to the heterotypic interaction [7]. Based on this theory, many investigators assumed of tumor and its microenvironment [1].