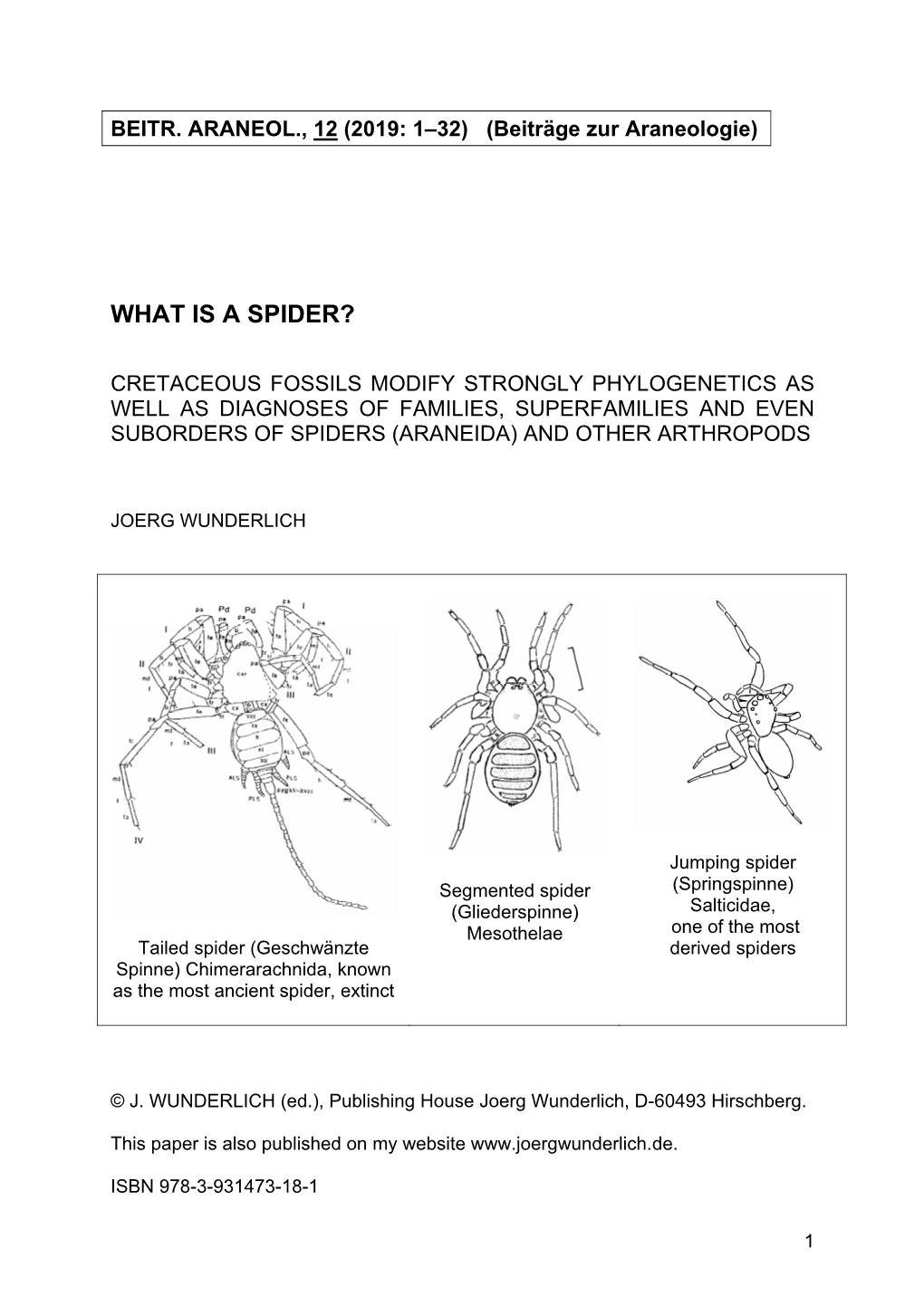
What Is a Spider?
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

The First Record of Family Segestriidae Simon, 1893 (Araneae: Dysderoidea) from Iran
Serket (2014) vol. 14(1): 15-18. The first record of family Segestriidae Simon, 1893 (Araneae: Dysderoidea) from Iran Alireza Zamani Department of Animal Biology, School of Biology and Center of Excellence in Phylogeny of Living Organisms in Iran, College of Science, University of Tehran, Tehran, Iran [email protected] Abstract The family Segestriidae Simon, 1893 and the species Segestria senoculata (Linnaeus, 1758) are recorded in Iran for the first time, based on a single female specimen. Keywords: Spiders, Segestriidae, Segestria senoculata, new record, Iran. Introduction Segestriidae Simon, 1893 is a small family of medium-sized, araneomorph, ecribellate, haplogyne spiders with three tarsal claws which are globally represented by 119 species in three genera (Platnick, 2014). These spiders are six-eyed, and are usually distinguishable by having their third pair of legs directed forwards. From taxonomic point of view, Segestriidae is closely related to Dysderidae, and are considered as a member of the superfamily Dysderoidea. The type genus, Segestria Latreille, 1804, is consisted of 18 species and one subspecies which are mostly distributed in the Palaearctic ecozone (Platnick, 2014). One of the more distributed species is Segestria senoculata (Linnaeus, 1758). This species, like most segestriids, occupies a wide variety of habitats; they prefer living in holes within walls and barks, or under stones, where they build a tubular retreat, with strong threads of silk radiating from the entrance (Roberts, 1995). So far, about 500 spider species of more than 38 families have been reported from Iran (based on our upcoming work on the renewed checklist and the history of studies), but no documentation of the family Segestriidae has been reported from Iran (Mozaffarian & Marusik, 2001; Ghavami, 2006; Kashefi et al., 2013). -

Sexual Selection Research on Spiders: Progress and Biases
Biol. Rev. (2005), 80, pp. 363–385. f Cambridge Philosophical Society 363 doi:10.1017/S1464793104006700 Printed in the United Kingdom Sexual selection research on spiders: progress and biases Bernhard A. Huber* Zoological Research Institute and Museum Alexander Koenig, Adenauerallee 160, 53113 Bonn, Germany (Received 7 June 2004; revised 25 November 2004; accepted 29 November 2004) ABSTRACT The renaissance of interest in sexual selection during the last decades has fuelled an extraordinary increase of scientific papers on the subject in spiders. Research has focused both on the process of sexual selection itself, for example on the signals and various modalities involved, and on the patterns, that is the outcome of mate choice and competition depending on certain parameters. Sexual selection has most clearly been demonstrated in cases involving visual and acoustical signals but most spiders are myopic and mute, relying rather on vibrations, chemical and tactile stimuli. This review argues that research has been biased towards modalities that are relatively easily accessible to the human observer. Circumstantial and comparative evidence indicates that sexual selection working via substrate-borne vibrations and tactile as well as chemical stimuli may be common and widespread in spiders. Pattern-oriented research has focused on several phenomena for which spiders offer excellent model objects, like sexual size dimorphism, nuptial feeding, sexual cannibalism, and sperm competition. The accumulating evidence argues for a highly complex set of explanations for seemingly uniform patterns like size dimorphism and sexual cannibalism. Sexual selection appears involved as well as natural selection and mechanisms that are adaptive in other contexts only. Sperm competition has resulted in a plethora of morpho- logical and behavioural adaptations, and simplistic models like those linking reproductive morphology with behaviour and sperm priority patterns in a straightforward way are being replaced by complex models involving an array of parameters. -

SEXUAL CONFLICT in ARACHNIDS Ph.D
MASARYK UNIVERSITY FACULTY OF SCIENCE DEPARTMENT OF BOTANY AND ZOOLOGY SEXUAL CONFLICT IN ARACHNIDS Ph.D. Dissertation Lenka Sentenská Supervisor: prof. Mgr. Stanislav Pekár, Ph.D. Brno 2017 Bibliographic Entry Author Mgr. Lenka Sentenská Faculty of Science, Masaryk University Department of Botany and Zoology Title of Thesis: Sexual conflict in arachnids Degree programme: Biology Field of Study: Ecology Supervisor: prof. Mgr. Stanislav Pekár, Ph.D. Academic Year: 2016/2017 Number of Pages: 199 Keywords: Sexual selection; Spiders; Scorpions; Sexual cannibalism; Mating plugs; Genital morphology; Courtship Bibliografický záznam Autor: Mgr. Lenka Sentenská Přírodovědecká fakulta, Masarykova univerzita Ústav botaniky a zoologie Název práce: Konflikt mezi pohlavími u pavoukovců Studijní program: Biologie Studijní obor: Ekologie Vedoucí práce: prof. Mgr. Stanislav Pekár, Ph.D. Akademický rok: 2016/2017 Počet stran: 199 Klíčová slova: Pohlavní výběr; Pavouci; Štíři; Sexuální kanibalismus; Pohlavní zátky; Morfologie genitálií; Námluvy ABSTRACT Sexual conflict is pervasive in all sexually reproducing taxa and is especially intense in carnivorous species, because the interaction between males and females encompasses the danger of getting killed instead of mated. Carnivorous arachnids, such as spiders and scorpions, are notoriously known for their cannibalistic tendencies. Studies of the conflict between arachnid males and females focus mainly on spiders because of the frequent occurrence of sexual cannibalism and unique genital morphology of both sexes. The morphology, in combination with common polyandry, further promotes the sexual conflict in form of an intense sperm competition and male tactics to reduce or avoid it. Scorpion females usually mate only once per litter, but the conflict between sexes is also intense, as females can be very aggressive, and so males engage in complicated mating dances including various components considered to reduce female aggression and elicit her cooperation. -

Untangling the Web… Spiders in Arizona Fields! Ayman Mostafa, Lydia M
Untangling the Web… Spiders in Arizona Fields! Ayman Mostafa, Lydia M. Brown, Tim Vandervoet, Peter C. Ellsworth (University of Arizona), Vonny Barlow (University of California) & Steven E. Naranjo (USDA-ARS, ALARC) Spiders are beneficial inhabitants of agricultural fields because of Lygus nymph prey their important contributions to biological control of pest insects, consuming tons of small arthropods every year. Spiders eat anything they can catch, even prey larger than themselves. When they are abundant, they contribute to the control of many insect pests in A Arizona crop fields including whiteflies, Lygus bugs, fleahoppers, Leafhopper and lepidopteran larvae. Field studies in Arizona demonstrate that the prey B crab spider, Misumenops celer (Family Thomisidae, Fig. 1A, B) and Dictyna spider, Dictyna reticulata (Family Dictynidae, Fig. 1C, D) are common in Arizona cotton fields and can be influential predators. Unlike other spiders that spin webs to capture their food, crab spiders rely on stealth and surprise. They actively search plant surfaces, litter, and debris for prey. They hide in flowers or foliage and ambush their prey. Their common name derives from the fact that they look like and walk like crabs. Dictyna are small, brownish, web-making E spiders that trap whitefly adults and other insects in their webs (Fig. 1C). Examining their webs enables easy identification of what D species of whitefly are in the field (sweetpotato or banded-winged). C Jumping spiders (Family Salticidae, Fig. 1E) are generally less abundant in cotton fields but, like crab spiders, ambush their prey. They have stout bodies and long front legs adapted for jumping, as well as four pairs of eyes with one very large set in the middle of their face. -

Prey of the Jumping Spider Phidippus Johnsoni (Araneae : Salticidae)
Jackson, R. R . 1977 . Prey of the jumping spider Phidippus johnsoni (Araneae : Salticidae) . J. Arachnol. 5 :145-149 . PREY OF THE JUMPING SPIDER PHIDIPPUS JOHNSONI (ARANEAE : SALTICIDAE) Robert R. Jackson I Zoology Departmen t University of Californi a Berkeley, California 9472 0 ABSTRACT Field data indicate that P. johnsoni is an euryphagous predator, whose diet includes organisms (aphids, ants, opilionids) sometimes considered distasteful to spiders . Other spiders are preyed upon , including conspecifics. Prey size tends to be one quarter to three quarters the size of the predator . INTRODUCTION Since spiders are probably a dominant group of predators of insects (Bristowe, 1941 ; Riechert, 1974; Turnbull, 1973), there is considerable interest in their feeding ecology . Spiders have usually been considered to be euryphagous predators with a stabilizing , rather than regulative, effect on insect populations (Riechert, 1974) . However, informa- tion concerning the prey taken by particular spider species, in the field, is limited . Field studies by Edgar (1969, 1970), Robinson and Robinson (1970) and Turnbull (1960) are especially noteworthy . During the course of a study of the reproductive biology of Phidippus johnsoni (Peckham and Peckham) (Jackson, 1976), occasionally individuals of this species were found in the field holding prey in their chelicerae . Each prey discovered in this way i s listed in Table 1 . In addition, Ken Evans and Charles Griswold, who were familiar wit h this species, recorded observations of P. johnsoni with prey. (Their data are included in Table 1 .) These data came from a variety of habitats in western North America, most o f which have been described elsewhere (Jackson, 1976) . -

North American Spiders of the Genera Cybaeus and Cybaeina
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by The University of Utah: J. Willard Marriott Digital... BULLETIN OF THE UNIVERSITY OF UTAH Volume 23 December, 1932 No. 2 North American Spiders of the Genera Cybaeus and Cybaeina BY RALPH V. CHAMBERLIN and WILTON IVIE BIOLOGICAL SERIES, Vol. II, No. / - PUBLISHED BY THE UNIVERSITY OF UTAH SALT LAKE CITY THE UNIVERSITY PRESS UNIVERSITY OF UTAH SALT LAKE CITY A Review of the North American Spider of the Genera Cybaeus and Cybaeina By R a l p h V. C h a m b e r l i n a n d W i l t o n I v i e The frequency with which members of the Agelenid genus Cybaeus appeared in collections made by the authors in the mountainous and timbered sections of the Pacific coast region and the representations therein of various apparently undescribed species led to the preparation of this review of the known North American forms. One species hereto fore placed in Cybaeus is made the type of a new genus Cybaeina. Most of our species occur in the western states; and it is probable that fur ther collecting in this region will bring to light a considerable number of additional forms. The drawings accompanying the paper were made from specimens direct excepting in a few cases where material was not available. In these cases the drawings were copied from the figures published by the authors of the species concerned, as indicated hereafter in each such case, but these drawings were somewhat revised to conform with the general scheme of the other figures in order to facilitate comparison. -

A Summary List of Fossil Spiders
A summary list of fossil spiders compiled by Jason A. Dunlop (Berlin), David Penney (Manchester) & Denise Jekel (Berlin) Suggested citation: Dunlop, J. A., Penney, D. & Jekel, D. 2010. A summary list of fossil spiders. In Platnick, N. I. (ed.) The world spider catalog, version 10.5. American Museum of Natural History, online at http://research.amnh.org/entomology/spiders/catalog/index.html Last udated: 10.12.2009 INTRODUCTION Fossil spiders have not been fully cataloged since Bonnet’s Bibliographia Araneorum and are not included in the current Catalog. Since Bonnet’s time there has been considerable progress in our understanding of the spider fossil record and numerous new taxa have been described. As part of a larger project to catalog the diversity of fossil arachnids and their relatives, our aim here is to offer a summary list of the known fossil spiders in their current systematic position; as a first step towards the eventual goal of combining fossil and Recent data within a single arachnological resource. To integrate our data as smoothly as possible with standards used for living spiders, our list follows the names and sequence of families adopted in the Catalog. For this reason some of the family groupings proposed in Wunderlich’s (2004, 2008) monographs of amber and copal spiders are not reflected here, and we encourage the reader to consult these studies for details and alternative opinions. Extinct families have been inserted in the position which we hope best reflects their probable affinities. Genus and species names were compiled from established lists and cross-referenced against the primary literature. -

Tarantulas and Social Spiders
Tarantulas and Social Spiders: A Tale of Sex and Silk by Jonathan Bull BSc (Hons) MSc ICL Thesis Presented to the Institute of Biology of The University of Nottingham in Partial Fulfilment of the Requirements for the Degree of Doctor of Philosophy The University of Nottingham May 2012 DEDICATION To my parents… …because they both said to dedicate it to the other… I dedicate it to both ii ACKNOWLEDGEMENTS First and foremost I would like to thank my supervisor Dr Sara Goodacre for her guidance and support. I am also hugely endebted to Dr Keith Spriggs who became my mentor in the field of RNA and without whom my understanding of the field would have been but a fraction of what it is now. Particular thanks go to Professor John Brookfield, an expert in the field of biological statistics and data retrieval. Likewise with Dr Susan Liddell for her proteomics assistance, a truly remarkable individual on par with Professor Brookfield in being able to simplify even the most complex techniques and analyses. Finally, I would really like to thank Janet Beccaloni for her time and resources at the Natural History Museum, London, permitting me access to the collections therein; ten years on and still a delight. Finally, amongst the greats, Alexander ‘Sasha’ Kondrashov… a true inspiration. I would also like to express my gratitude to those who, although may not have directly contributed, should not be forgotten due to their continued assistance and considerate nature: Dr Chris Wade (five straight hours of help was not uncommon!), Sue Buxton (direct to my bench creepy crawlies), Sheila Keeble (ventures and cleans where others dare not), Alice Young (read/checked my thesis and overcame her arachnophobia!) and all those in the Centre for Biomolecular Sciences. -

Cribellum and Calamistrum Ontogeny in the Spider Family Uloboridae: Linking Functionally Related but Sbparate Silk Spinning Features
2OOl. The Journal of Arachnology 29:22O-226 CRIBELLUM AND CALAMISTRUM ONTOGENY IN THE SPIDER FAMILY ULOBORIDAE: LINKING FUNCTIONALLY RELATED BUT SBPARATE SILK SPINNING FEATURES Brent D. Opelt: Department of BioLogy, Virginia Polytechnic Institute and State University, Blacksburg, Virginia 24061 USA ABSTRACT. The fourth metatarsusof cribellatespiders bears a setal comb, the calamistrum,that sweeps over the cribellum, drawing fibrils from its spigots and helping to combine these with the capture thread's supporting fibers. In four uloborid species (Hyptiotes cavatus, Miagrammopes animotus, Octonoba sinensis, Uloborus glomosus), calamistrum length and cribellum width have similar developmental trajec- tories, despite being borne on different regions of the body. In contrast, developmental rates of metatarsus IV and its calamistrum differ within species and vary independently among species. Thus, the growth rates of metatarsus IV and the calamistrum are not coupled, freeing calamistrum length to track cribellum width and metatarsus IV length to respond to changes in such features as combing behavior and abdomen dimensions. Keywords: Cribellar thread, Hyptiotes cavatus, Miagrammopes animotus, Octonoba sinensis, Ulobortts glomosus Members of the family Uloboridae produce ond instars (Opell 1979). However, their cri- cribellar prey capture threads formed of a bella and calamistra are not functional until sheath of fine, looped fibrils that surround par- they molt again to become third instars. Sec- acribellar and axial supporting fibers (Eber- ond instar orb-weaving uloborid species pro- hard & Pereira 1993: Opell 1990, 1994, 1995, duce a juvenile web that lacks a sticky spiral 1996, 1999: Peters 1983, 1984, 1986). Cri- and has many closely spaced radii (Lubin bellar fibrils come from the spigots of an oval 1986). -

Causes and Consequences of External Female Genital Mutilation
Causes and consequences of external female genital mutilation I n a u g u r a l d i s s e r t a t i o n Zur Erlangung des akademischen Grades eines Doktors der Naturwissenschaften (Dr. rer. Nat.) der Mathematisch-Naturwissenschaftlichen Fakultät der Universität Greifswald Vorgelegt von Pierick Mouginot Greifswald, 14.12.2018 Dekan: Prof. Dr. Werner Weitschies 1. Gutachter: Prof. Dr. Gabriele Uhl 2. Gutachter: Prof. Dr. Klaus Reinhardt Datum der Promotion: 13.03.2019 Contents Abstract ................................................................................................................................................. 5 1. Introduction ................................................................................................................................... 6 1.1. Background ............................................................................................................................. 6 1.2. Aims of the presented work ................................................................................................ 14 2. References ................................................................................................................................... 16 3. Publications .................................................................................................................................. 22 3.1. Chapter 1: Securing paternity by mutilating female genitalia in spiders .......................... 23 3.2. Chapter 2: Evolution of external female genital mutilation: why do males harm their mates?.................................................................................................................................. -

Taxonomic Notes on Amaurobius (Araneae: Amaurobiidae), Including the Description of a New Species
Zootaxa 4718 (1): 047–056 ISSN 1175-5326 (print edition) https://www.mapress.com/j/zt/ Article ZOOTAXA Copyright © 2020 Magnolia Press ISSN 1175-5334 (online edition) https://doi.org/10.11646/zootaxa.4718.1.3 http://zoobank.org/urn:lsid:zoobank.org:pub:5F484F4E-28C2-44E4-B646-58CBF375C4C9 Taxonomic notes on Amaurobius (Araneae: Amaurobiidae), including the description of a new species YURI M. MARUSIK1,2, S. OTTO3 & G. JAPOSHVILI4,5 1Institute for Biological Problems of the North RAS, Portovaya Str. 18, Magadan, Russia. E-mail: [email protected] 2Department of Zoology & Entomology, University of the Free State, Bloemfontein 9300, South Africa 3GutsMuthsstr. 42, 04177 Leipzig, Germany. 4Institute of Entomology, Agricultural University of Georgia, Agmashenebeli Alley 13 km, 0159 Tbilisi, Georgia 5Invertebrate Research Center, Tetrtsklebi, Telavi municipality 2200, Georgia 6Corresponding author. E-mail: [email protected] Abstract A new species, Amaurobius caucasicus sp. n., is described based on the holotype male and two male paratypes from Eastern Georgia. A similar species, A. hercegovinensis Kulczyński, 1915, known only from the original description is redescribed. The taxonomic status of Amaurobius species considered as nomina dubia and species described outside the Holarctic are also assessed. Amaurobius koponeni Marusik, Ballarin & Omelko, 2012, syn. n. described from northern India is a junior synonym of A. jugorum L. Koch, 1868 and Amaurobius yanoianus Nakatsudi, 1943, syn. n. described from Micronesia is synonymised with the titanoecid species Pandava laminata (Thorell, 1878) a species known from Eastern Africa to Polynesia. Considerable size variation in A. antipovae Marusik et Kovblyuk, 2004 is briefly discussed. Key words: Aranei, Asia, Caucasus, Georgia, Kakheti, misplaced, new synonym, nomen dubium, redescription Introduction Amaurobius C.L. -

Amaurobiidae, Macrobuninae)
Zootaxa 3931 (3): 387–400 ISSN 1175-5326 (print edition) www.mapress.com/zootaxa/ Article ZOOTAXA Copyright © 2015 Magnolia Press ISSN 1175-5334 (online edition) http://dx.doi.org/10.11646/zootaxa.3931.3.3 http://zoobank.org/urn:lsid:zoobank.org:pub:13BA1A35-C31A-4910-8EAC-1D840C5CAD18 Revision of Emmenomma Simon (Amaurobiidae, Macrobuninae) ALMEIDA-SILVA LINA M.1,2,3, GRISWOLD CHARLES E.1 & BRESCOVIT ANTONIO D.2 1California Academy of Sciences, Department of Entomology, 55 Music Concourse Drive, Golden Gate Park, 94118, San Francisco, USA. E-mail: [email protected], [email protected] 2Laboratório Especial de Coleções Zoológicas - LECZ, Instituto Butantan, Avenida Vital Brazil, 1500, 05503-900, São Paulo, Brazil. E-mail: [email protected], [email protected] 3Corresponding author Abstract The genus Emmenomma is revised and now includes three species from Southern Chile, Argentina and Falkland Islands (Islas Malvinas). The type species Emmenomma oculatum is redescribed and considered a senior synonym of E. beauchenicum. Emmenomma oculatum obscurum is removed from synonymy with E. oculatum, raised to the species level and redescribed; the male of this species remains unknown. A new species, Emmenomma joshuabelli sp. nov. is described. The presence of a grate shaped tapetum outside the Lycosoidea clade is described. Detailed images are provided for all known species. Key words: Chile, Spiders, Naevius, Anisacate, Tibial gland Introduction The spider genus Emmenomma was proposed by Simon (1884) based on several male and female specimens from Tierra del Fuego, Chile. Due to its atypical eye pattern, with greatly enlarged lateral eyes, and peculiar male palp, with an elbowed and flexibly attached dorsal tibial apophysis (DTA) that rests on the retrolateral side of the palp and can be easily mistaken for a RTA, Emmenomma is not easily confused with other spiders and there are no synonyms of the generic name.