Research Article Review Jmb
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Heterologous Expression and Identification of the Genes Involved
Heterologous Expression and Identification of the Genes Involved in Anaerobic Degradation of 1,3-Dihydroxybenzene (Resorcinol) in Azoarcus anaerobiusᰔ Paula I. Darley,1† Jutta A. Hellstern,1†‡ Javier I. Medina-Bellver,2 Silvia Marque´s,2 Bernhard Schink,1 and Bodo Philipp1* Fachbereich Biologie, Universita¨t Konstanz, D-78457 Constance, Germany,1 and Estacio´n Experimental del Zaidı´n, C/. Profesor Albareda 1, E-18008 Granada, Spain2 Received 9 November 2006/Accepted 9 March 2007 Azoarcus anaerobius, a strictly anaerobic, gram-negative bacterium, utilizes resorcinol as a sole carbon and energy source with nitrate as an electron acceptor. Previously, we showed that resorcinol degradation by this bacterium is initiated by two oxidative steps, both catalyzed by membrane-associated enzymes that lead to the formation of hydroxyhydroquinone (HHQ; 1,2,4-benzenetriol) and 2-hydroxy-1,4-benzoquinone (HBQ). This study presents evidence for the further degradation of HBQ in cell extracts to form acetic and malic acids. To identify the A. anaerobius genes required for anaerobic resorcinol catabolism, a cosmid library with genomic DNA was constructed and transformed into the phylogenetically related species Thauera aromatica, which cannot grow with resorcinol. By heterologous complementation, a transconjugant was identified that gained the ability to metabolize resorcinol. Its cosmid, designated R؉, carries a 29.88-kb chromosomal DNA fragment containing 22 putative genes. In cell extracts of T. aromatica transconjugants, resorcinol was degraded to HHQ, HBQ, and acetate, suggesting that cosmid R؉ carried all of the genes necessary for resorcinol degradation. On the basis of the physiological characterization of T. aromatica transconjugants carrying transposon insertions -in different genes of cosmid R؉, eight open reading frames were found to be essential for resorcinol miner alization. -

Metaproteogenomic Insights Beyond Bacterial Response to Naphthalene
ORIGINAL ARTICLE ISME Journal – Original article Metaproteogenomic insights beyond bacterial response to 5 naphthalene exposure and bio-stimulation María-Eugenia Guazzaroni, Florian-Alexander Herbst, Iván Lores, Javier Tamames, Ana Isabel Peláez, Nieves López-Cortés, María Alcaide, Mercedes V. del Pozo, José María Vieites, Martin von Bergen, José Luis R. Gallego, Rafael Bargiela, Arantxa López-López, Dietmar H. Pieper, Ramón Rosselló-Móra, Jesús Sánchez, Jana Seifert and Manuel Ferrer 10 Supporting Online Material includes Text (Supporting Materials and Methods) Tables S1 to S9 Figures S1 to S7 1 SUPPORTING TEXT Supporting Materials and Methods Soil characterisation Soil pH was measured in a suspension of soil and water (1:2.5) with a glass electrode, and 5 electrical conductivity was measured in the same extract (diluted 1:5). Primary soil characteristics were determined using standard techniques, such as dichromate oxidation (organic matter content), the Kjeldahl method (nitrogen content), the Olsen method (phosphorus content) and a Bernard calcimeter (carbonate content). The Bouyoucos Densimetry method was used to establish textural data. Exchangeable cations (Ca, Mg, K and 10 Na) extracted with 1 M NH 4Cl and exchangeable aluminium extracted with 1 M KCl were determined using atomic absorption/emission spectrophotometry with an AA200 PerkinElmer analyser. The effective cation exchange capacity (ECEC) was calculated as the sum of the values of the last two measurements (sum of the exchangeable cations and the exchangeable Al). Analyses were performed immediately after sampling. 15 Hydrocarbon analysis Extraction (5 g of sample N and Nbs) was performed with dichloromethane:acetone (1:1) using a Soxtherm extraction apparatus (Gerhardt GmbH & Co. -

University of Birmingham Synthesis of -Glucan in Mycobacteria Involves A
University of Birmingham Synthesis of -glucan in mycobacteria involves a hetero-octameric complex of trehalose synthase TreS and Maltokinase Pep2 Roy, Rana; Veeraraghavan, Usha; Kermani, Ali; Scott, David J; Hyde, Eva I; Besra, Gurdyal S; Alderwick, Luke J; Fütterer, Klaus DOI: 10.1021/cb400508k License: Creative Commons: Attribution (CC BY) Document Version Publisher's PDF, also known as Version of record Citation for published version (Harvard): Roy, R, Veeraraghavan, U, Kermani, A, Scott, DJ, Hyde, EI, Besra, GS, Alderwick, LJ & Fütterer, K 2013, 'Synthesis of -glucan in mycobacteria involves a hetero-octameric complex of trehalose synthase TreS and Maltokinase Pep2', ACS chemical biology, vol. 8, no. 10, pp. 2245-2255. https://doi.org/10.1021/cb400508k Link to publication on Research at Birmingham portal Publisher Rights Statement: Eligibility for repository : checked 30/06/2014 General rights Unless a licence is specified above, all rights (including copyright and moral rights) in this document are retained by the authors and/or the copyright holders. The express permission of the copyright holder must be obtained for any use of this material other than for purposes permitted by law. •Users may freely distribute the URL that is used to identify this publication. •Users may download and/or print one copy of the publication from the University of Birmingham research portal for the purpose of private study or non-commercial research. •User may use extracts from the document in line with the concept of ‘fair dealing’ under the Copyright, Designs and Patents Act 1988 (?) •Users may not further distribute the material nor use it for the purposes of commercial gain. -

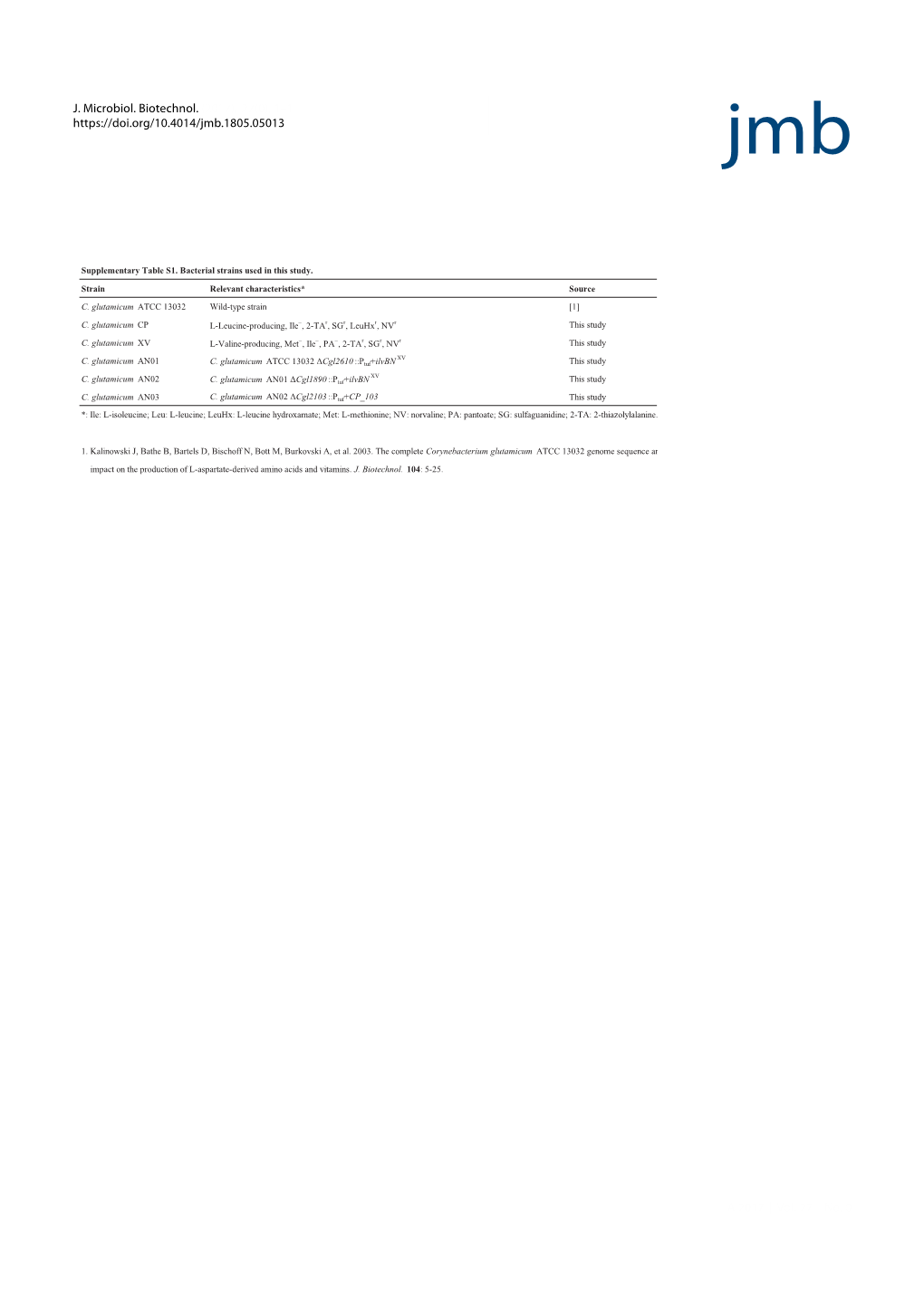
Table S1-Final.Xlsx
Table S1. Functional gene families covered on the GeoChip 5.0. No. of sequence‐ No. of group‐ No. of total No. of covered No. of total Category Subcategory Subcategory 2 Gene Encoded Enzyme specific probesa specific probesa probes on 5M CDSa probes on 5S Categories for Geochemical Cycling Carbon Cycling Carbon Degradation Agar beta_agarase Agarase 42 79 217 121 Agar Total 42 79 0 217 121 Alginate alginase Alginase 45 200 512 245 Alginate Total 45 200 0 512 245 Cellulose Endoglucanase Endoglucanase 198 248 446 740 343 Cellulose Total 198 248 446 740 343 Chitin Acetylglucosaminidase Acetylglucosaminidase 119 976 1095 3088 1116 Chitinase Chitinase 649 1231 1880 3285 1411 endochitinase Endochitinase 213 334 1104 547 exochitinase Exochitinase 14 55 259 69 Chitin Total 995 2596 2975 7736 3143 Glyoxylate cycle AceA Isocitrate lyase 69 373 442 887 0 AceB Malate synthase A 90 610 700 1457 0 Glyoxylate cycle Total 159 983 1142 2344 0 Hemicellulose Ara Arabinofuranosidase 172 543 715 1367 827 Mannanase Mannanase 159 238 397 639 478 Xylanase Xylanase 150 653 803 1739 858 Hemicellulose Total 481 1434 1915 3745 2163 Heparin heparinase Heparinase 7 53 166 60 Heparin Total 7 53 0 166 60 Hyaluronic acid hyaluronidase Hyaluronidase 880 383 88 Hyaluronidase Total 8 80 0 383 88 Lignin Glx Glyoxal oxidase 103 40 143 178 125 Mnp Manganese peroxidase 50 13 63 74 66 Phenol_oxidase Laccase or phenol oxidase 188 372 560 1009 677 Lignin Total 341 425 766 1261 868 Pectin Pectinase (pectate_lyase) Pectate lyase 53 252 305 827 321 Pme Pectin methylesterase 25 240 265 -

Cultural Soil
Structural and functional metage- nomic analyses of a tropical agri- cultural soil Análisis metagenómicos estructurales y funcionales de un suelo agrícola tropical Análises metagenómicas estruturais e funcionais de um solo agrícola tropical AUTHORS Received: 04.07.2018 Revised: 09.12.2018 Accepted: 21.01.2019 Lateef Babatunde 1 Salam@, 1 ABSTRACT babssalaam@yahoo. com Understanding the intricate link between the soil microbiota and their metabolic functions is important for agricultural and ecological processes and could be used as a biomarker of soil health. Oluwafemi Sunday To understand the relationship between soil microbial community structure and functions, a soil 2 Obayori microcosm designated 2S (agricultural soil) was set up. Metagenomic DNA was extracted from the soil microcosm and sequenced using Miseq Illumina next generation sequencing and analysed for their structural and functional properties. Structural analysis of the soil microcosm by MG-RAST @ Corresponding Author revealed 40 phyla, 78 classes, 157 orders, 273 families and 750 genera. Actinobacteria (54.0%) and 1Department of Biological Proteobacteria (17.5%) are the dominant phyla while Conexibacter (8.38%), Thermoleophilum (7.40%), Sciences, Al-Hikmah and Streptomyces (4.14%) are the dominant genera. Further assignment of the metagenomics using University, Ilorin, Kwara State, Nigeria. Cluster of Orthologous Groups (COG), Kyoto Encyclopedia of Genes and Genomes (KEGG), GhostKOALA, and NCBI’s CDD revealed diverse metabolic pathways utilized by the microbial 2Department of Microbiology, Lagos State community for the metabolism of carbohydrates, amino acids, lipids, biosynthesis of secondary University, Ojo, Lagos metabolites and resistance to antibiotics. Taxonomic analysis of the annotated genes also revealed State, Nigeria. the preponderance of members of Actinobacteria and Proteobacteria. -

Maleylacetate Reductase from Trichosporon Cutaneum
Biochem. J. (1980) 185, 783-786 783 Printed in Great Britain Maleylacetate Reductase from Trichosporon cutaneum Andras B. GAAL and Halina Y. NEUJAHR Department ofBiochemistry and Biochemical Technology, The Royal Institute of Technology, S 100 44 Stockholm, Sweden (Received 12 November 1979) The enzyme catalysing the reduction of maleylacetate to 3-oxoadipate was purified 150-fold from Trichosporon cutaneum, induced for aromatic metabolism by growth with resorcinol as a major carbon source. The enzyme separated upon electrofocusing into three species with pI values 4.6, 5.1 and 5.6. They had similar catalytic properties and the same molecular weight. Our group has previously reported that the yeast fer, pH 7.6, 0.2pmol of maleylacetate, 0.2,mol of Trichosporon cutaneum can be induced to metabol- NADPH and 50-200#ug of enzyme protein. Under ize phenol and resorcinol, both phenols being these conditions 1 enzyme unit corresponds to an attacked by the same hydroxylating enzyme and the absorbance decrease of 6.1 A340 units/min. same ring-cleavage enzyme (Neujahr & Varga, Determination of molecular weight. Molecular 1970; Varga & Neujahr, 1970). We have since weight was determined by gel filtration (Andrews, demonstrated that both phenol and resorcinol are 1965) on a Sephadex G-150 column metabolized to 3-oxoadipate (Gaal & Neujahr, (2.5cm x 85cm) equilibrated with 50mM-Tris/ 1979). The metabolism of resorcinol goes through H2SO4 buffer, pH 7.6. Alcohol dehydrogenase 1,2,4-trihydroxybenzene and maleylacetate. The (mol.wt. 150000), lactate dehydrogenase (mol.wt. enzyme catalysing the reduction of maleylacetate to 136000), hexokinase (mol.wt. 105000) and avidin 3-oxoadipate was enriched from crude extracts (mol.wt. -

Biochemical Studies on Redox Regulation in Different Dormancy Models of Mycobacteria
Biochemical Studies on Redox Regulation in Different Dormancy Models of Mycobacteria Thesis Submitted to Savitribai Phule Pune University For The Degree Of DOCTOR OF PHILOSOPHY IN BIOTECHNOLOGY By Ketaki Dilip Shurpali Research Supervisor Dr. Dhiman Sarkar Combichem Bioresource Center Organic Chemistry Division CSIR-National Chemical Laboratory Pune-411008 India October 2014 Certificate This is to certify that the work incorporated in the thesis entitled “ Biochemical Studies on Redox Regulation in Different Dormancy Models of Mycobacteria ” submitted by Ketaki Dilip Shurpali was carried out under my supervision at Combichem Bioresource Center, Organic Chemistry Division, National Chemical Laboratory, Pune-411008, Maharashtra, India. Materials obtained from other sources have been duly acknowledged in the thesis. Dr. Dhiman Sarkar (Research Guide) I Declaration by Research Scholar I hereby declare that the thesis entitled " Biochemical Studies on Redox Regulation in Different Dormancy Models of Mycobacteria ", submitted for the Degree of Doctor of Philosophy to the Savitribai Phule Pune University, has been carried out by me at Combichem Bioresource Center, Organic Chemistry Division, CSIR-National Chemical Laboratory, Pune-411008, Maharashtra, India, under the supervision of Dr. Dhiman Sarkar (Research supervisor). The work is original and has not been submitted in part or full by me for any other degree or diploma to any other University. Ketaki Dilip Shurpali (Research Scholar) II Dedications This thesis is dedicated to all those people who were always besides me in my good and bad times. First dedication goes to my father Mr. Dilip Shurpali and mother Mrs. Devyani Shurpali who instilled in me the desire to learn new things and confidence to achieve my dreams. -

Bacterial Degradation of Chlorophenols and Their Derivatives Pankaj Kumar Arora and Hanhong Bae*
Arora and Bae Microbial Cell Factories 2014, 13:31 http://www.microbialcellfactories.com/content/13/1/31 REVIEW Open Access Bacterial degradation of chlorophenols and their derivatives Pankaj Kumar Arora and Hanhong Bae* Abstract Chlorophenols (CPs) and their derivatives are persistent environmental pollutants which are used in the manufacture of dyes, drugs, pesticides and other industrial products. CPs, which include monochlorophenols, polychlorophenols, chloronitrophenols, chloroaminophenols and chloromethylphenols, are highly toxic to living beings due to their carcinogenic, mutagenic and cytotoxic properties. Several physico-chemical and biological methods have been used for removal of CPs from the environment. Bacterial degradation has been considered a cost-effective and eco-friendly method of removing CPs from the environment. Several bacteria that use CPs as their sole carbon and energy sources have been isolated and characterized. Additionally, the metabolic pathways for degradation of CPs have been studied in bacteria and the genes and enzymes involved in the degradation of various CPs have been identified and characterized. This review describes the biochemical and genetic basis of the degradation of CPs and their derivatives. Keywords: Chlorophenol, Environmental pollutants, Bacterial degradation, Biodegradation Introduction human carcinogens [5]. Similarly, the United States Envir- Chlorophenols (CPs) are aromatic ring structures contain- onmental Protection Agency has included several CPs in its ing at least one chlorine atom (−Cl) and one hydroxyl list of priority pollutants. (−OH) group at the benzene rings. Five groups of CPs have Several conventional methods such as adsorption, ion been recognized on the basis of their chemical structures, exchange, liquid–liquid extraction, and chemical oxida- monochlorophenols (MCPs), polychlorophenols (poly- tion and advanced oxidation processes have been used CPs), chloronitrophenols (CNPs), chloroaminophenols for the removal of CPs from wastewater [3,6]. -

United States Patent (10) Patent No.: US 9,562,241 B2 Burk Et Al
USOO9562241 B2 (12) United States Patent (10) Patent No.: US 9,562,241 B2 Burk et al. (45) Date of Patent: Feb. 7, 2017 (54) SEMI-SYNTHETIC TEREPHTHALIC ACID 5,487.987 A * 1/1996 Frost .................... C12N 9,0069 VLAMCROORGANISMIS THAT PRODUCE 5,504.004 A 4/1996 Guettler et all 435,142 MUCONCACID 5,521,075- W I A 5/1996 Guettler et al.a (71) Applicant: GENOMATICA, INC., San Diego, CA 3. A '95 seal. (US) 5,686,276 A 11/1997 Lafend et al. 5,700.934 A 12/1997 Wolters et al. (72) Inventors: Mark J. Burk, San Diego, CA (US); (Continued) Robin E. Osterhout, San Diego, CA (US); Jun Sun, San Diego, CA (US) FOREIGN PATENT DOCUMENTS (73) Assignee: Genomatica, Inc., San Diego, CA (US) CN 1 358 841 T 2002 EP O 494 O78 7, 1992 (*) Notice: Subject to any disclaimer, the term of this (Continued) patent is extended or adjusted under 35 U.S.C. 154(b) by 0 days. OTHER PUBLICATIONS (21) Appl. No.: 14/308,292 Abadjieva et al., “The Yeast ARG7 Gene Product is Autoproteolyzed to Two Subunit Peptides, Yielding Active (22) Filed: Jun. 18, 2014 Ornithine Acetyltransferase,” J. Biol. Chem. 275(15): 11361-11367 2000). (65) Prior Publication Data . al., “Discovery of amide (peptide) bond synthetic activity in US 2014/0302573 A1 Oct. 9, 2014 Acyl-CoA synthetase,” J. Biol. Chem. 28.3(17): 11312-11321 (2008). Aberhart and Hsu, "Stereospecific hydrogen loss in the conversion Related U.S. Application Data of H, isobutyrate to f—hydroxyisobutyrate in Pseudomonas (63) Continuation of application No. -

Evidence of Two Oxidative Reaction Steps Initiating Anaerobic Degradation of Resorcinol (1,3-Dihydroxybenzene) by the Denitrifying Bacterium Azoarcus Anaerobius
JOURNAL OF BACTERIOLOGY, July 1998, p. 3644–3649 Vol. 180, No. 14 0021-9193/98/$04.0010 Copyright © 1998, American Society for Microbiology. All Rights Reserved. Evidence of Two Oxidative Reaction Steps Initiating Anaerobic Degradation of Resorcinol (1,3-Dihydroxybenzene) by the Denitrifying Bacterium Azoarcus anaerobius BODO PHILIPP* AND BERNHARD SCHINK Fakulta¨t fu¨r Biologie, Mikrobielle O¨ kologie, Universita¨t Konstanz, D-78457 Konstanz, Germany Received 21 January 1998/Accepted 11 May 1998 The denitrifying bacterium Azoarcus anaerobius LuFRes1 grows anaerobically with resorcinol (1,3-dihydroxy- benzene) as the sole source of carbon and energy. The anaerobic degradation of this compound was investi- gated in cell extracts. Resorcinol reductase, the key enzyme for resorcinol catabolism in fermenting bacteria, was not present in this organism. Instead, resorcinol was hydroxylated to hydroxyhydroquinone (HHQ; 1,2,4-trihydroxybenzene) with nitrate or K3Fe(CN)6 as the electron acceptor. HHQ was further oxidized with nitrate to 2-hydroxy-1,4-benzoquinone as identified by high-pressure liquid chromatography, UV/visible light spectroscopy, and mass spectroscopy. Average specific activities were 60 mU mg of protein21 for resorcinol hydroxylation and 150 mU mg of protein21 for HHQ dehydrogenation. Both activities were found nearly exclusively in the membrane fraction and were only barely detectable in extracts of cells grown with benzoate, indicating that both reactions were specific for resorcinol degradation. These findings suggest a new strategy of anaerobic degradation of aromatic compounds involving oxidative steps for destabilization of the aromatic ring, different from the reductive dearomatization mechanisms described so far. The biochemistry of anaerobic degradation of aromatic In this work, we reexamined resorcinol degradation by A. -

Masterarbeit / Master's Thesis
MASTERARBEIT / MASTER’S THESIS Titel der Masterarbeit / Title of the Master‘s Thesis „Physiology, Metabolism and Biohydrogen Production of Desulfurococcus fermentans“ verfasst von / submitted by Barbara Reischl, BSc angestrebter akademischer Grad / in partial fulfilment of the requirements for the degree of Master of Science (MSc) Wien, 2016 / Vienna 2016 Studienkennzahl lt. Studienblatt / A 066 830 degree programme code as it appears on the student record sheet: Studienrichtung lt. Studienblatt / Molecular Microbiology, Microbial Ecology degree programme as it appears on and Immunobiology the student record sheet: Betreut von / Supervisor: Univ.-Prof. Dipl.-Biol. Dr. Christa Schleper Mitbetreut von / Co-Supervisor: Mag. Mag. Dr. Simon Karl-Maria Rasso Rittmann, Bakk. 2 Table of Contents 1. Introduction .......................................................................................................... 5 1.1 Role of Biohydrogen ................................................................................................ 5 1.2 Hydrogenases ......................................................................................................... 6 1.3 Characteristics of Desulfurococcus fermentans ....................................................... 7 1.4 Aims of this Study ................................................................................................... 8 2. Material and Methods ............................................................................................ 9 2.1 Chemicals .............................................................................................................. -

Glucan in Mycobacteria Involves a Hetero-Octameric Complex of Trehalose Synthase Tres and Maltokinase Pep2
Synthesis of -glucan in mycobacteria involves a hetero-octameric complex of trehalose synthase TreS and Maltokinase Pep2 Roy, Rana; Veeraraghavan, Usha; Kermani, Ali; Scott, David; Hyde, Eva; Besra, Gurdyal; Alderwick, Luke; Futterer, Klaus DOI: 10.1021/cb400508k License: Creative Commons: Attribution (CC BY) Document Version Publisher's PDF, also known as Version of record Citation for published version (Harvard): Roy, R, Veeraraghavan, U, Kermani, A, Scott, DJ, Hyde, EI, Besra, GS, Alderwick, LJ & Fütterer, K 2013, 'Synthesis of -glucan in mycobacteria involves a hetero-octameric complex of trehalose synthase TreS and Maltokinase Pep2', ACS chemical biology, vol. 8, no. 10, pp. 2245-2255. https://doi.org/10.1021/cb400508k Link to publication on Research at Birmingham portal Publisher Rights Statement: Eligibility for repository : checked 30/06/2014 General rights Unless a licence is specified above, all rights (including copyright and moral rights) in this document are retained by the authors and/or the copyright holders. The express permission of the copyright holder must be obtained for any use of this material other than for purposes permitted by law. •Users may freely distribute the URL that is used to identify this publication. •Users may download and/or print one copy of the publication from the University of Birmingham research portal for the purpose of private study or non-commercial research. •User may use extracts from the document in line with the concept of ‘fair dealing’ under the Copyright, Designs and Patents Act 1988 (?) •Users may not further distribute the material nor use it for the purposes of commercial gain. Where a licence is displayed above, please note the terms and conditions of the licence govern your use of this document.