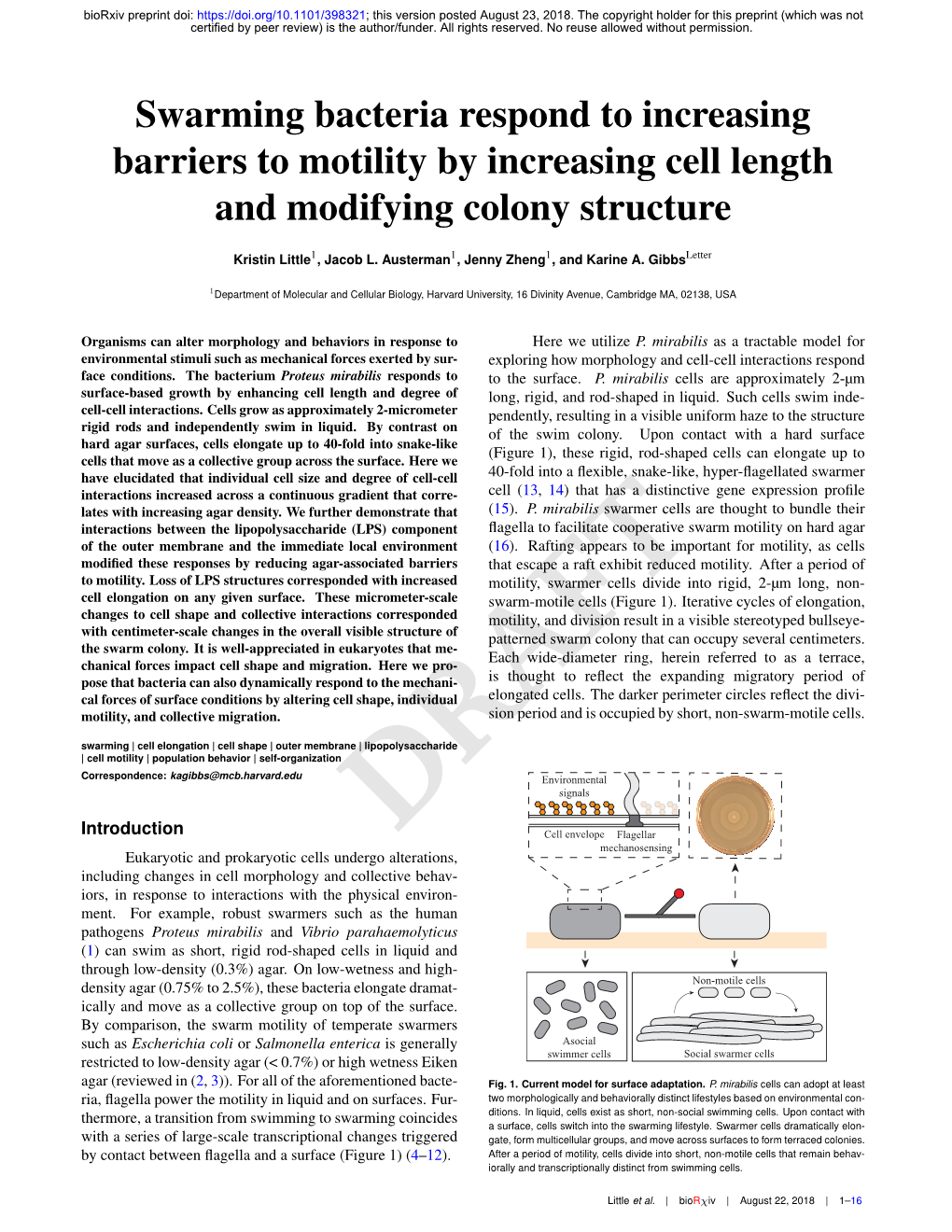
Swarming Bacteria Respond to Increasing Barriers to Motility by Increasing Cell Length and Modifying Colony Structure
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Pulse Generation in the Quorum Machinery of Pseudomonas Aeruginosa
This is a repository copy of Pulse generation in the quorum machinery of Pseudomonas aeruginosa. White Rose Research Online URL for this paper: https://eprints.whiterose.ac.uk/116745/ Version: Published Version Article: Alfiniyah, Cicik Alfiniyah, Bees, Martin Alan and Wood, Andrew James orcid.org/0000- 0002-6119-852X (2017) Pulse generation in the quorum machinery of Pseudomonas aeruginosa. Bulletin of Mathematical Biology. pp. 1360-1389. ISSN 1522-9602 https://doi.org/10.1007/s11538-017-0288-z Reuse This article is distributed under the terms of the Creative Commons Attribution (CC BY) licence. This licence allows you to distribute, remix, tweak, and build upon the work, even commercially, as long as you credit the authors for the original work. More information and the full terms of the licence here: https://creativecommons.org/licenses/ Takedown If you consider content in White Rose Research Online to be in breach of UK law, please notify us by emailing [email protected] including the URL of the record and the reason for the withdrawal request. [email protected] https://eprints.whiterose.ac.uk/ Bull Math Biol (2017) 79:1360–1389 DOI 10.1007/s11538-017-0288-z ORIGINAL ARTICLE Pulse Generation in the Quorum Machinery of Pseudomonas aeruginosa Cicik Alfiniyah1,2 · Martin A. Bees1 · A. Jamie Wood1,3 Received: 28 September 2016 / Accepted: 3 May 2017 / Published online: 19 May 2017 © The Author(s) 2017. This article is an open access publication Abstract Pseudomonas aeruginosa is a Gram-negative bacterium that is responsible for a wide range of infections in humans. -

Subtilase Sprp Exerts Pleiotropic Effects in Pseudomonas Aeruginosa
ORIGINAL RESEARCH Subtilase SprP exerts pleiotropic effects in Pseudomonas aeruginosa Alexander Pelzer1, Tino Polen2, Horst Funken1, Frank Rosenau3, Susanne Wilhelm1, Michael Bott2 & Karl-Erich Jaeger1 1Institute of Molecular Enzyme Technology, Research Centre Juelich, Heinrich-Heine-University Duesseldorf, D-52426 Juelich, Germany 2Institut of Bio- und Geosciences IBG-1: Biotechnology, Research Centre Juelich, D-52426 Juelich, Germany 3Institute of Pharmaceutical Biotechnology, Ulm-University, Albert-Einstein-Allee 11, D-89069 Ulm, Germany Keywords Abstract Biofilm, microarray, motility, orf PA1242, protease, Pseudomonas aeruginosa, The open reading frame PA1242 in the genome of Pseudomonas aeruginosa Pyoverdine. PAO1 encodes a putative protease belonging to the peptidase S8 family of sub- tilases. The respective enzyme termed SprP consists of an N-terminal signal Correspondence peptide and a so-called S8 domain linked by a domain of unknown function Karl-Erich Jaeger, Institute of Molecular (DUF). Presumably, this DUF domain defines a discrete class of Pseudomonas Enzyme Technology, Heinrich-Heine- proteins as homologous domains can be identified almost exclusively in pro- University Duesseldorf, Research Centre Juelich, D-52426 Juelich, Germany. teins of the genus Pseudomonas. The sprP gene was expressed in Escherichia coli Δ Tel: (+49)-2461-61-3716; Fax: (+49)-2461-61- and proteolytic activity was demonstrated. A P. aeruginosa sprP mutant was 2490; E-mail: [email protected] constructed and its gene expression pattern compared to the wild-type strain by genome microarray analysis revealing altered expression levels of 218 genes. Funding Information Apparently, SprP is involved in regulation of a variety of different cellular pro- 2021 As a member of the CLIB -Graduate cesses in P. -

Social Behaviour in Microorganisms 13
13 Social behaviour in microorganisms Kevin R. Foster OVERVIEW Sociobiology has come a long way. We now have Many species do all of this in surface-attached a solid base of evolutionary theory supported communities, known as biofilms, in which the by a myriad of empirical tests. It is perhaps less diversity of species and interactions reaches appreciated, however, that first discussions of bewildering heights. Grouping can even involve social behaviour and evolution in Darwin’s day differentiation and development, as in the spec- drew upon single-celled organisms. Since then, tacular multicellular escape responses of slime microbes have received short shrift, and their moulds and myxobacteria. Like any society, how- full spectrum of sociality has only recently come ever, microbes face conflict, and most groups to light. Almost everything that a microorgan- will involve instances of both cooperation and ism does has social consequences; simply divid- competition among their members. And, as in ing can consume another’s resources. Microbes any society, microbial conflicts are mediated by also secrete a wide range of products that affect three key processes: constraints on rebellion, others, including digestive enzymes, toxins, mol- coercion that enforces compliance, and kinship ecules for communication and DNA that allows whereby cells direct altruistic aid towards clone- genes to mix both within and among species. mates. 13.1 Introduction indeed sociology, microbes have featured in descrip- tions of social life. Prominent among these are the We must be prepared to learn some day, from the students writings of Herbert Spencer, the social philosopher of microscopical pond-life, facts of unconscious mutual sup- who coined the term survival of the fi ttest in the wake port, even from the life of micro-organisms. -

Ecological and Evolutionary Implications of Shape During Population Expansion
ECOLOGICAL AND EVOLUTIONARY IMPLICATIONS OF SHAPE DURING POPULATION EXPANSION Submitted by Christopher Lee Coles to the University of Exeter as a thesis for the degree of Doctor of Philosophy in Biological Sciences January 2015 This thesis is available for library use on the understanding that it is copyright material and that no quotation from the thesis may be published without proper acknowledgement. I certify that all material in this thesis which is not my own work has been identified and that no material has previously been submitted and approved for the award of a degree by this or any other University. Signature: ................................... ~ 1 ~ In loving memory of Laura Joan Coles ~ 2 ~ Abstract The spatial spread of populations is one of the most visible and fundamental processes in population and community ecology. Due to the potential negative impacts of spatial spread of invasive populations, there has been intensive research into understanding the drivers of ecological spread, predicting spatial dynamics, and finding management strategies that best constrain or control population expansion. However, understanding the spread of populations has proved to be a formidable task and our ability to accurately predict the spread of these populations has to date been limited. Microbial populations, during their spread across agar plate environments, can exhibit a wide array of spatial patterns, ranging from relatively circular patterns to highly irregular, fractal-like patterns. Work analysing these patterns of spread has mainly focused on the underlying mechanistic processes responsible for these patterns, with relatively little investigation into the ecological and evolutionary drivers of these patterns. With the increased recognition of the links between microbial and macrobial species, it is possible that many of the ecological/evolutionary mechanisms responsible for these patterns of spread at a microbial level extrapolate to the spatial spread of populations in general. -

A Preliminary Study of Chemical Profiles of Honey, Cerumen
foods Article A Preliminary Study of Chemical Profiles of Honey, Cerumen, and Propolis of the African Stingless Bee Meliponula ferruginea Milena Popova 1 , Dessislava Gerginova 1 , Boryana Trusheva 1 , Svetlana Simova 1, Alfred Ngenge Tamfu 2 , Ozgur Ceylan 3, Kerry Clark 4 and Vassya Bankova 1,* 1 Institute of Organic Chemistry with Centre of Phytochemistry, Bulgarian Academy of Sciences, 1113 Sofia, Bulgaria; [email protected] (M.P.); [email protected] (D.G.); [email protected] (B.T.); [email protected] (S.S.) 2 Department of Chemical Engineering, School of Chemical Engineering and Mineral Industries, University of Ngaoundere, 454 Ngaoundere, Cameroon; [email protected] 3 Food Quality Control and Analysis Program, Ula Ali Kocman Vocational School, Mugla Sitki Kocman University, 48147 Ula Mugla, Turkey; [email protected] 4 Volunteer Advisor in Beekeeping and Bee Products with Canadian Executive Services Organization, P.O. Box 2090, Dawson Creek, BC V1G 4K8, Canada; [email protected] * Correspondence: [email protected]; Tel.: +359-2-9606-149 Abstract: Recently, the honey and propolis of stingless bees have been attracting growing atten- tion because of their health-promoting properties. However, studies on these products of African Meliponini are still very scarce. In this preliminary study, we analyzed the chemical composition of honey, two cerumen, and two resin deposits (propolis) samples of Meliponula ferruginea from Tanzania. The honey of M. ferruginea was profiled by NMR and indicated different long-term stability Citation: Popova, M.; Gerginova, D.; from Apis mellifera European (Bulgarian) honey. It differed significantly in sugar and organic acids Trusheva, B.; Simova, S.; Tamfu, A.N.; content and had a very high amount of the disaccharide trehalulose, known for its bioactivities. -

Cell Density and Mobility Protect Swarming Bacteria Against Antibiotics
Cell density and mobility protect swarming bacteria against antibiotics Mitchell T. Butler, Qingfeng Wang, and Rasika M. Harshey1 Section of Molecular Genetics and Microbiology, and Institute of Cellular and Molecular Biology, University of Texas, Austin, TX 78712 Edited by Raghavendra Gadagkar, Indian Institute of Science, Bangalore, India, and approved December 9, 2009 (received for review September 23, 2009) Swarming bacteria move in multicellular groups and exhibit Bacillus and Serratia—and show that cell density and mobility are adaptive resistance to multiple antibiotics. Analysis of this phe- common protective features for survival against antimicrobials. nomenon has revealed the protective power of high cell densities to withstand exposure to otherwise lethal antibiotic concentra- Results and Discussion tions. We find that high densities promote bacterial survival, even Antibiotic Resistance Is a Property of High Cell Density and Is Favored in a nonswarming state, but that the ability to move, as well as the by Mobility. In soft agar (0.3%), cells swim individually inside the speed of movement, confers an added advantage, making swarm- agar and are referred to as swimmers. In medium agar (0.6%), ing an effective strategy for prevailing against antimicrobials. We cells move as a group on the surface and are referred to as find no evidence of induced resistance pathways or quorum- swarmers. The original E-test strip assay showing differential sensing mechanisms controlling this group resistance, which occurs antibiotic resistance of swimmer and swarmer cells of Salmonella at a cost to cells directly exposed to the antibiotic. This work has (13, 14) is shown in Fig. 1A. -

Learning the Space-Time Phase Diagram of Bacterial Swarm Expansion
Learning the space-time phase diagram of bacterial swarm expansion Hannah Jeckela,b,c, Eric Jellia,b, Raimo Hartmanna, Praveen K. Singha, Rachel Mokc,d, Jan Frederik Totze, Lucia Vidakovica, Bruno Eckhardtb,Jorn¨ Dunkelc,1, and Knut Dreschera,b,1 aMax Planck Institute for Terrestrial Microbiology, 35043 Marburg, Germany; bFachbereich Physik and LOEWE Zentrum fur¨ Synthetische Mikrobiologie SYNMIKRO, Philipps-Universitat¨ Marburg, 35032 Marburg, Germany; cDepartment of Mathematics, Massachusetts Institute of Technology, Cambridge, MA 02139; dDepartment of Mechanical Engineering, Massachusetts Institute of Technology, Cambridge, MA 02139; and eInstitute for Theoretical Physics, Technische Universitat¨ Berlin, 10623 Berlin, Germany Edited by David A. Weitz, Harvard University, Cambridge, MA, and approved December 11, 2018 (received for review July 7, 2018) Coordinated dynamics of individual components in active matter Results and Discussion are an essential aspect of life on all scales. Establishing a com- To track the swarming behavior of B. subtilis over five orders of prehensive, causal connection between intracellular, intercellular, magnitude in space at the single-cell level, we developed an adap- and macroscopic behaviors has remained a major challenge due tive microscope that acquires high-speed movies at times and to limitations in data acquisition and analysis techniques suit- locations determined by a live feedback between image feature able for multiscale dynamics. Here, we combine a high-through- recognition and an automated movement of the scanning area put adaptive microscopy approach with machine learning, to (Fig. 1A). This technique allows us to image a radially expanding identify key biological and physical mechanisms that determine swarm at single-cell resolution in space and time (Fig. -
![Spatial Awareness of a Bacterial Swarm Harshitha S. Kotian[1]](https://docslib.b-cdn.net/cover/3201/spatial-awareness-of-a-bacterial-swarm-harshitha-s-kotian-1-2003201.webp)
Spatial Awareness of a Bacterial Swarm Harshitha S. Kotian[1]
Spatial Awareness of a Bacterial Swarm Harshitha S. Kotian[1][2], Shalini Harkar[2], Shubham Joge[3], Ayushi Mishra[3], Amith Zafal[1], Varsha Singh[3], Manoj M. Varma[1][2] [1]Centre for Nano Science and Engineering, Indian Institute of Science, Bangalore, India [2] Robert Bosch Centre for Cyber Physical Systems, Indian Institute of Science, Bangalore, India [3]Molecular Reproduction, Development and Genetics, Indian Institute of Science, Bangalore, India Abstract Bacteria are perhaps the simplest living systems capable of complex behaviour involving sensing and coherent, collective behaviour an example of which is the phenomena of swarming on agar surfaces. Two fundamental questions in bacterial swarming is how the information gathered by individual members of the swarm is shared across the swarm leading to coordinated swarm behaviour and what specific advantages does membership of the swarm provide its members in learning about their environment. In this article, we show a remarkable example of the collective advantage of a bacterial swarm which enables it to sense inert obstacles along its path. Agent based computational model of swarming revealed that independent individual behaviour in response to a two-component signalling mechanism could produce such behaviour. This is striking because independent individual behaviour without any explicit communication between agents was found to be sufficient for the swarm to effectively compute the gradient of signalling molecule concentration across the swarm and respond to it. Introduction Bacteria are perhaps the simplest living systems capable of complex behaviour involving sensing and coherent, collective behaviour an example of which is the phenomena of swarming, where bacteria colonize a solid surface (typically, nutrient loaded agar) in geometric patterns characteristic of different species (Ben-Jacob, 1997; Kearns, 2010). -

Motility, Mixing, and Multicellularity
Genet Program Evolvable Mach (2007) 8:115–129 DOI 10.1007/s10710-007-9029-7 ORIGINAL PAPER Motility, mixing, and multicellularity Cristian A. Solari Æ John O. Kessler Æ Raymond E. Goldstein Received: 30 April 2006 / Revised: 19 February 2007 / Published online: 10 May 2007 Ó Springer Science+Business Media, LLC 2007 Abstract A fundamental issue in evolutionary biology is the transition from unicellular to multicellular organisms, and the cellular differentiation that accom- panies the increase in group size. Here we consider recent results on two types of ‘‘multicellular’’ systems, one produced by many unicellular organisms acting collectively, and another that is permanently multicellular. The former system is represented by groups of the bacterium Bacillus subtilis and the latter is represented by members of the colonial volvocalean green algae. In these flagellated organisms, the biology of chemotaxis, metabolism and cell–cell signaling is intimately con- nected to the physics of buoyancy, motility, diffusion, and mixing. Our results include the discovery in bacterial suspensions of intermittent episodes of disorder and collective coherence characterized by transient, recurring vortex streets and high-speed jets of cooperative swimming. These flow structures markedly enhance transport of passive tracers, and therefore likely have significant implications for intercellular communication. Experiments on the Volvocales reveal that the sterile flagellated somatic cells arrayed on the surface of Volvox colonies are not only important for allowing motion toward light (phototaxis), but also play a crucial role C. A. Solari (&) Department of Ecology and Evolutionary Biology, University of Arizona, Tucson, AZ 85721, USA e-mail: [email protected] J. O. -

Dead Cells Release a 'Necrosignal' That Activates Antibiotic Survival Pathways in Bacterial Swarms
ARTICLE https://doi.org/10.1038/s41467-020-17709-0 OPEN Dead cells release a ‘necrosignal’ that activates antibiotic survival pathways in bacterial swarms ✉ Souvik Bhattacharyya 1, David M. Walker 1 & Rasika M. Harshey 1 Swarming is a form of collective bacterial motion enabled by flagella on the surface of semi- solid media. Swarming populations exhibit non-genetic or adaptive resistance to antibiotics, despite sustaining considerable cell death. Here, we show that antibiotic-induced death of a fi 1234567890():,; sub-population bene ts the swarm by enhancing adaptive resistance in the surviving cells. Killed cells release a resistance-enhancing factor that we identify as AcrA, a periplasmic component of RND efflux pumps. The released AcrA interacts on the surface of live cells with an outer membrane component of the efflux pump, TolC, stimulating drug efflux and inducing expression of other efflux pumps. This phenomenon, which we call ‘necrosignaling’, exists in other Gram-negative and Gram-positive bacteria and displays species-specificity. Given that adaptive resistance is a known incubator for evolving genetic resistance, our findings might be clinically relevant to the rise of multidrug resistance. 1 Department of Molecular Biosciences, University of Texas at Austin, Austin, TX 78712, USA. ✉email: [email protected] NATURE COMMUNICATIONS | (2020) 11:4157 | https://doi.org/10.1038/s41467-020-17709-0 | www.nature.com/naturecommunications 1 ARTICLE NATURE COMMUNICATIONS | https://doi.org/10.1038/s41467-020-17709-0 acteria employ many appendages for movement and dis- reversed at both higher Kan concentrations as well as later times, Bpersal in their ecological niches1. Of these, flagella-driven where planktonic cells became more susceptible. -

LIIS ANDRESEN Regulation of Virulence in Plant-Pathogenic
DISSERTATIONES BIOLOGICAE LIIS ANDRESEN LIIS UNIVERSITATIS TARTUENSIS 228 Regulation of virulence in plant-pathogenic pectobacteria LIIS ANDRESEN Regulation of virulence in plant-pathogenic pectobacteria Tartu 2012 ISSN 1024–6479 ISBN 978–9949–32–146–9 DISSERTATIONES BIOLOGICAE UNIVERSITATIS TARTUENSIS 229 DISSERTATIONES BIOLOGICAE UNIVERSITATIS TARTUENSIS 229 LIIS ANDRESEN Regulation of virulence in plant-pathogenic pectobacteria Institute of Molecular and Cell Biology, University of Tartu, Estonia Dissertation is accepted for the commencement of the degree of Doctor of Philosophy (in Gene Technology) on 26.09.2012 by the Council of the Institute of Molecular and Cell Biology, University of Tartu Supervisors: Dotsent Andres Mäe, PhD Institute of Molecular and Cell Biology University of Tartu Estonia Opponent: Guy Condemine, PhD National Institute for Applied Sciences, Lyon National Center for Scientific Research France Commencement: Room No 105, Institute of Molecular and Cell Biology, University of Tartu; 23b Riia Str., Tartu, on November 16th, 2012, at 10.00 The University of Tartu grants the publication of this dissertation. ISSN 1024–6479 ISBN 978–9949–32–146–9 (print) ISBN 978–9949–32–147–6 (pdf) Copyright: Liis Andresen, 2012 University of Tartu Press www.tyk.ee Order No. 510 TABLE OF CONTENTS LIST OF ORIGINAL PUBLICATIONS ....................................................... 6 ABBREVIATIONS ........................................................................................ 7 INTRODUCTION ......................................................................................... -

Model Socialite, Problem Pathogen: the Evolution and Ecology of Cooperation in the Bacterium Pseudomonas Aeruginosa
Model socialite, problem pathogen: the evolution and ecology of cooperation in the bacterium Pseudomonas aeruginosa Adin Ross-Gillespie Thesis submitted for the degree of Doctor of Philosophy University of Edinburgh 2008 2 "The most important unanswered question in evolutionary biology, and more generally in the social sciences, is how cooperative behaviour evolved and can be maintained in human or other animal groups and societies.” - Lord Robert May, former Chief Scientific Adviser to HM Government, in his final presidential address to the Royal Society, 30 November 2005. 3 4 Abstract In recent decades we have learned that cooperation is an important and pervasive feature of microbial life. This revelation raises exciting possibilities. On the one hand, we can now augment our understanding of how social phenomena evolve by using microbial model systems to test our theories. On the other hand, we can use concepts from social evolution to gain insight into the biology of the microbes we hope to control or kill. In this thesis I explore both possibilities. First, I consider the theoretical problem of how and when microbial cooperation might be subject to frequency- and density- dependence. Formerly, vague theory and a scant, sometimes contradictory empirical literature made it unclear when such patterns could be expected. Here, I develop theory tailored to a microbial context, and in each case, I test key predictions from the theory in laboratory experiments, using as my model trait the production of siderophores by the bacterium Pseudomonas aeruginosa . Secondly, I consider the ecological consequences of cooperator-cheat dynamics in the context of an infection.