Controversies on the Origin of Life
Total Page:16
File Type:pdf, Size:1020Kb

Load more
Recommended publications
-

BWTB Nov. 13Th Dukes 2016
1 Playlist Nov. 13th 2016 LIVE! From DUKES in Malibu 9AM / OPEN Three hours non stop uninterrupted Music from JPG&R…as we broadcast LIVE from DUKES in Malibu…. John Lennon – Steel and Glass - Walls And Bridges ‘74 Much like “How Do You Sleep” three years earlier, this is another blistering Lennon track that sets its sights on Allen Klein (who had contributed lyrics to “How Do You Sleep” those few years before). The Beatles - Revolution 1 - The Beatles 2 The first song recorded during the sessions for the “White Album.” At the time of its recording, this slower version was the only version of John Lennon’s “Revolution,” and it carried that titled without a “1” or a “9” in the title. Recording began on May 30, 1968, and 18 takes were recorded. On the final take, the first with a lead vocal, the song continued past the 4 1/2 minute mark and went onto an extended jam. It would end at 10:17 with John shouting to the others and to the control room “OK, I’ve had enough!” The final six minutes were pure chaos with discordant instrumental jamming, plenty of feedback, percussive clicks (which are heard in the song’s introduction as well), and John repeatedly screaming “alright” and moaning along with his girlfriend, Yoko Ono. Ono also spoke random streams of consciousness on the track such as “if you become naked.” This bizarre six-minute section was clipped off the version of what would become “Revolution 1” to form the basis of “Revolution 9.” Yoko’s “naked” line appears in the released version of “Revolution 9” at 7:53. -

John Lennon from ‘Imagine’ to Martyrdom Paul Mccartney Wings – Band on the Run George Harrison All Things Must Pass Ringo Starr the Boogaloo Beatle
THE YEARS 1970 -19 8 0 John Lennon From ‘Imagine’ to martyrdom Paul McCartney Wings – band on the run George Harrison All things must pass Ringo Starr The boogaloo Beatle The genuine article VOLUME 2 ISSUE 3 UK £5.99 Packed with classic interviews, reviews and photos from the archives of NME and Melody Maker www.jackdaniels.com ©2005 Jack Daniel’s. All Rights Reserved. JACK DANIEL’S and OLD NO. 7 are registered trademarks. A fine sippin’ whiskey is best enjoyed responsibly. by Billy Preston t’s hard to believe it’s been over sent word for me to come by, we got to – all I remember was we had a groove going and 40 years since I fi rst met The jamming and one thing led to another and someone said “take a solo”, then when the album Beatles in Hamburg in 1962. I ended up recording in the studio with came out my name was there on the song. Plenty I arrived to do a two-week them. The press called me the Fifth Beatle of other musicians worked with them at that time, residency at the Star Club with but I was just really happy to be there. people like Eric Clapton, but they chose to give me Little Richard. He was a hero of theirs Things were hard for them then, Brian a credit for which I’m very grateful. so they were in awe and I think they had died and there was a lot of politics I ended up signing to Apple and making were impressed with me too because and money hassles with Apple, but we a couple of albums with them and in turn had I was only 16 and holding down a job got on personality-wise and they grew to the opportunity to work on their solo albums. -

The History of Planet Earth
SECOND EDITION Earth’s Evolving The History of Systems Planet Earth Ronald Martin, Ph.D. University of Delaware Newark, Delaware © Jones & Bartlett Learning, LLC, an Ascend Learning Company. NOT FOR SALE OR DISTRIBUTION 9781284457162_FMxx_00i_xxii.indd 1 07/11/16 1:46 pm World Headquarters Jones & Bartlett Learning 5 Wall Street Burlington, MA 01803 978-443-5000 [email protected] www.jblearning.com Jones & Bartlett Learning books and products are available through most bookstores and online booksellers. To contact Jones & Bartlett Learning directly, call 800-832-0034, fax 978-443-8000, or visit our website, www.jblearning.com. Substantial discounts on bulk quantities of Jones & Bartlett Learning publications are available to corporations, professional associations, and other qualified organizations. For details and specific discount information, contact the special sales department at Jones & Bartlett Learning via the above contact information or send an email to [email protected]. Copyright © 2018 by Jones & Bartlett Learning, LLC, an Ascend Learning Company All rights reserved. No part of the material protected by this copyright may be reproduced or utilized in any form, electronic or mechanical, including photocopying, recording, or by any information storage and retrieval system, without written permission from the copyright owner. The content, statements, views, and opinions herein are the sole expression of the respective authors and not that of Jones & Bartlett Learning, LLC. Reference herein to any specific commercial product, process, or service by trade name, trademark, manufacturer, or otherwise does not constitute or imply its endorsement or recommendation by Jones & Bartlett Learning, LLC, and such reference shall not be used for advertis- ing or product endorsement purposes. -

Finding Life Beyond Earth
FINDING LIFE BEYOND EARTH Education Collection NOVA Education pbs.org/nova/education, [email protected] ! NOVA’s “Finding Life Beyond Earth” script visualized. NOVA Education pbs.org/nova/education, [email protected] ! Table of Contents Credits and Acknowledgements! 2 Overview! 4 1 — Meet the Planets! 13 2 — What Is Life?! 16 3 — Basic Ingredients for Life! 19 4 — Extreme Living! 22 5 — Home Sweet Home! 25 6 — Where to Look for Life! 29 7 — How to Search! 33 Common Misconceptions! 38 Related Resources! 39 Appendices! 40 © 2012 WGBH Educational Foundation!!1 Credits and Acknowledgements Produced by NOVA’s Department of Education, WGBH Rachel Connolly, Director of Education | NOVA Chris Randall, Senior Editorial Project Director | WGBH Education Department Maiken Lilley, Education Coordinator | NOVA Graham Veth, Education Outreach Coordinator | NOVA Rachel Gesserman, Production Assistant | NOVA Advisors Chris Dietlin, WGBH Kristen Erickson, NASA Irena Fayngold, WGBH Kay Ferrari, NASA/JPL Rebecca Jaramillo, National Institute of Aerospace Daniella Scalice, NASA Astrobiology Institute Harla Sherwood, National Institute of Aerospace Stephanie Shipp, Lunar and Planetary Institute Anita Sohus, NASA/JPL “Finding Life Beyond Earth” is produced in cooperation with NASA and the National Institute of Aerospace. Additional funding is provided by Millicent Bell through the Millicent and Eugene Bell Foundation. This material is based on work supported by the National Aeronautics and Space Administration, Langley Research Center, under the Research Cooperative Agreement No. NNL09AA00A awarded to the National Institute of Aerospace. Any opinions, findings, and conclusions or recommendations expressed in this publication are those of WGBH and do not necessarily reflect the views of the National Institute of Aerospace. -

Nature Based Solutions FEMA
PROMOTING NATURE-BASED HAZARD MITIGATION THROUGH FEMA MITIGATION GRANTS ABBREVIATIONS ADCIRC – Advanced Circulation Model HGM Approach – Hydrogeomorphic Approach BCA – Benefit-Cost Analysis HMA – Hazard Mitigation Assistance BCR – Benefit-Cost Ratio HMGP – Hazard Mitigation Grant Program BRIC – Building Resilient Infrastructure and MSCP – Multiple Species Conservation Program Communities NBS – Nature-Based Solution C&CB – Capability- and Capacity-Building NFIP – National Flood Insurance Program CDBG-DR – Community Development Block Grant- Disaster Recovery NFWF – National Fish and Wildlife Foundation CDBG-MIT – Community Development Block NOAA – National Oceanic and Atmospheric Grant-Mitigation Administration D.C. – District of Columbia NOFO – Notice of Funding Opportunity DEM – Department of Emergency Management NPV – Net Present Value DOI – Department of the Interior SCC – State Coastal Conservancy EDYS – Ecological Dynamics Simulation SDG&E – San Diego Gas & Electric EMA – Emergency Management Agency SFHA – Special Flood Hazard Area EPA SWMM – Environmental Protection Agency SHMO – State Hazard Mitigation Officer Storm Water Management Model SLAMM – Sea Level Affecting Marshes Model FEMA – Federal Emergency Management Agency SRH-2D – Sedimentation and River Hydraulics – FIRM – Flood Insurance Rate Map Two-Dimension FMA – Flood Mitigation Assistance STWAVE – Steady-State Spectral Wave Model FMAG – Fire Management Assistance Grant TNC – The Nature Conservancy HAZUS – Hazards US USACE – U.S. Army Corps of Engineers HEC-HMS – Hydrologic -

Download This Article in PDF Format
BIO Web of Conferences 4, 00014 (2015) DOI: 10.1051/bioconf/20150400014 C Owned by the authors, published by EDP Sciences, 2015 From neonts to filionts and their progenies... Biodiversity draws its origins from the origins of life itself Marie-Christine Maurela Institut de Systématique, Évolution, Biodiversité (ISYEB - UMR 7205 – CNRS, MNHN, UPMC, EPHE), Muséum national d’Histoire naturelle, Sorbonne Universités, 45 rue Buffon, CP. 50, 75005 Paris, France “Should the question of the priority of the egg over the chicken, or of the chicken over the egg disturb you, it is because you assume that animals were originally what they are to-day; what madness!" The dream of d’Alembert, 1760, Denis Diderot Abstract. Since the origins, different modes of life exist. Indeed it is difficult to imagine how things could have been constructed by a single event that would have marked the birth of life. A brief historical background of the subject will be presented here. Then, we shall wander through the history of the discoveries that have led the way of experimental science over the last 150 years and shall dwell on one of the major paradigms, that of the RNA World. This paradigm illustrates the path followed by the living that tries all possible means when expressed at the molecular level as simple free molecule leading to RNA fragments and to modern subviral particles such as viroids then to ramifications and compartments. Between the ancient and the modern RNA world we underline how environment talks to molecules leading to the global living tissus which establish the ground of biodiversity. -

The Solar System
5 The Solar System R. Lynne Jones, Steven R. Chesley, Paul A. Abell, Michael E. Brown, Josef Durech,ˇ Yanga R. Fern´andez,Alan W. Harris, Matt J. Holman, Zeljkoˇ Ivezi´c,R. Jedicke, Mikko Kaasalainen, Nathan A. Kaib, Zoran Kneˇzevi´c,Andrea Milani, Alex Parker, Stephen T. Ridgway, David E. Trilling, Bojan Vrˇsnak LSST will provide huge advances in our knowledge of millions of astronomical objects “close to home’”– the small bodies in our Solar System. Previous studies of these small bodies have led to dramatic changes in our understanding of the process of planet formation and evolution, and the relationship between our Solar System and other systems. Beyond providing asteroid targets for space missions or igniting popular interest in observing a new comet or learning about a new distant icy dwarf planet, these small bodies also serve as large populations of “test particles,” recording the dynamical history of the giant planets, revealing the nature of the Solar System impactor population over time, and illustrating the size distributions of planetesimals, which were the building blocks of planets. In this chapter, a brief introduction to the different populations of small bodies in the Solar System (§ 5.1) is followed by a summary of the number of objects of each population that LSST is expected to find (§ 5.2). Some of the Solar System science that LSST will address is presented through the rest of the chapter, starting with the insights into planetary formation and evolution gained through the small body population orbital distributions (§ 5.3). The effects of collisional evolution in the Main Belt and Kuiper Belt are discussed in the next two sections, along with the implications for the determination of the size distribution in the Main Belt (§ 5.4) and possibilities for identifying wide binaries and understanding the environment in the early outer Solar System in § 5.5. -

George Porter
G EORGE P O R T E R Flash photolysis and some of its applications Nobel Lecture, December 11, 1967 One of the principal activities of man as scientist and technologist has been the extension of the very limited senses with which he is endowed so as to enable him to observe phenomena with dimensions very different from those he can normally experience. In the realm of the very small, microscopes and micro- balances have permitted him to observe things which have smaller extension or mass than he can see or feel. In the dimension of time, without the aid of special techniques, he is limited in his perception to times between about one twentieth of a second ( the response time of the eye) and about 2·10 9 seconds (his lifetime). Y et most of the fundamental processes and events, particularly those in the molecular world which we call chemistry, occur in milliseconds or less and it is therefore natural that the chemist should seek methods for the study of events in microtime. My own work on "the study of extremely fast chemical reactions effected by disturbing the equilibrium by means of very short pulses of energy" was begun in Cambridge twenty years ago. In 1947 I attended a discussion of the Faraday Society on "The Labile Molecule". Although this meeting was en- tirely concerned with studies of short lived chemical substances, the four hundred pages of printed discussion contain little or no indication of the im- pending change in experimental approach which was to result from the intro- . -
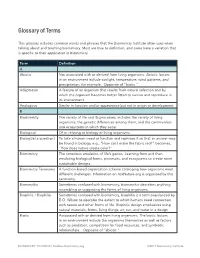
Glossary of Terms
Glossary of Terms This glossary includes common words and phrases that the Biomimicry Institute often uses when talking about and teaching biomimicry. Most are true to definition, and some have a variation that is specific to their application in biomimicry. Term Definition A Abiotic Not associated with or derived from living organisms. Abiotic factors in an environment include sunlight, temperature, wind patterns, and precipitation, for example. Opposite of "biotic." Adaptation A feature of an organism that results from natural selection and by which the organism becomes better fitted to survive and reproduce in its environment. Analogous Similar in function and/or appearance but not in origin or development. B Biodiversity The variety of life and its processes; includes the variety of living organisms, the genetic differences among them, and the communities and ecosystems in which they occur. Biological Of or relating to biology or living organisms. Biologize [a question] To take a human need or function and rephrase it so that an answer may be found in biology, e.g., "How can I make the fabric red?" becomes, "How does nature create color?" Biomimicry The conscious emulation of life’s genius. Learning from and then emulating biological forms, processes, and ecosystems to create more sustainable designs. Biomimicry Taxonomy A function-based organization scheme cataloging how organisms meet different challenges. Information on AskNature.org is organized by this taxonomy. Biomorphic Sometimes confused with biomimicry, biomorphic describes anything resembling or suggesting the forms of living organisms. Biophilic / Biophilia Sometimes confused with biomimicry, biophilia is a term popularized by E.O. Wilson to describe the extent to which humans need connection with nature and other forms of life. -
Manfred Eigen: the Realization of His Vision of Biophysical Chemistry
CORE Metadata, citation and similar papers at core.ac.uk Provided by OIST Institutional Repository Manfred Eigen: the realization of his vision of Biophysical Chemistry Author Herbert Jackle, Carmen Rotte, Peter Gruss journal or European Biophysics Journal publication title volume 47 number 4 page range 319-323 year 2017-12-11 Publisher Springer International Publishing Rights (C) 2017 The Author(s). Author's flag publisher URL http://id.nii.ac.jp/1394/00000696/ doi: info:doi/10.1007/s00249-017-1266-y Creative Commons Attribution 4.0 International (http://creativecommons.org/licenses/by/4.0/) European Biophysics Journal (2018) 47:319–323 https://doi.org/10.1007/s00249-017-1266-y REVIEW Manfred Eigen: the realization of his vision of Biophysical Chemistry Herbert Jäckle1 · Carmen Rotte1 · Peter Gruss1,2 Received: 27 August 2017 / Accepted: 11 November 2017 / Published online: 11 December 2017 © The Author(s) 2017. This article is an open access publication Abstract Manfred Eigen turned 90 on May 9th, 2017. He celebrated with a small group of colleagues and friends on behalf of the many inspired by him over his lifetime—whether scientists, artists, or philosophers. A small group of friends, because many—who by their breakthroughs have changed the face of science in diferent research areas—have already died. But it was a special day, devoted to the many genius facets of Manfred Eigen’s oeuvre, and a day to highlight the way in which he continues to exude a great, vital and unbroken passion for science as well as an insatiable curiosity beyond his own scientifc interests. -

Leslie E. Orgel 1927–2007
Leslie E. Orgel 1927–2007 A Biographical Memoir by Jack D. Dunitz and Gerald F. Joyce ©2013 National Academy of Sciences. Any opinions expressed in this memoir are those of the authors and do not necessarily reflect the views of the National Academy of Sciences. LESLIE ELEAZER ORGEL January 12, 1927–October 27, 2007 Elected to the NAS, 1990 Leslie Eleazer Orgel was a theoretical chemist and inves- tigator of the origins of life who made deep and lasting contributions in both of these scientific areas. He was born in London, England, on January 12, 1927, the second of three children of Simon and Deborah (Gnivisch) Orgel. His older brother Nevill was born on July 2, 1922, and died on December 28, 1957. His younger sister Delia was born on June 19, 1933, and currently resides in Silver Spring, Maryland. Leslie Orgel died on October 27, 2007, in San Diego, California, from pancreatic cancer. He is survived by his wife of 57 years, Alice (Levinson) Orgel; by his three children, Vivienne (b. April 4, 1955), Richard (b. November 29, 1956), and Robert (b. June 25, 1968); and by five By Jack D. Dunitz grandchildren. and Gerald F. Joyce After attending Dame Alice Owen’s School in London, which was evacuated during World War II to Bedford, England, Orgel studied chemistry at the University of Oxford, graduating in 1948 as BA with First Class Honours in Chem- istry. He then undertook graduate research with Leslie Sutton, senior chemistry tutor at Magdalen College and himself a distinguished physical chemist. Orgel’s1 first publication (1951) dealt with the semi-empirical calculation of electric dipole moments of conjugated heterocyclic molecules, and can be of no more than historical interest today. -

Henry Taube by J
Chemical Education Today Nobel Centennial Essays A Century of Chemical Dynamics Traced through the Nobel Prizes W 1983: Henry Taube by J. Van Houten Nobel Prize in Chemistry 1983 Photo by Rudy Baum Henry Taube (1915– ) for his work on the mechanisms of electron transfer Henry Taube, shown on reactions, especially in metal complexes the cover of C&E News. This is the eighth in a series of essays (1) written in com- Reprinted with permis- memoration of the centennial of the Nobel Prize, examin- sion from Chemical & ing the history of chemical dynamics in the 20th century.W Engineering News, May 21, 1984. © As his Nobel citation (2) states, Henry Taube studied elec- Copyright 1984 Ameri- tron transfer reactions of transition metal complexes. The can Chemical Society. Nobel Prize to Taube came exactly seventy years after it was awarded to Alfred Werner (3) for developing the structural chemistry of inorganic transition metal complexes. During the entire 20th century, Taube and Werner are the only in- to Werner’s. His early mechanistic studies relied heavily on organic transition metal chemists to win a Nobel Prize.1 In isotope tracer techniques developed by George de Hevesy, the fact, many of the complexes that Taube studied were first 1943 Nobel Laureate (1d). The second of the 100 references prepared or characterized by Werner. in Taube’s Nobel lecture (6) is to a 1920 paper where de Taube’s citation (2) concludes with the statement: “There Hevesy used naturally occurring lead isotopes to follow the is no doubt that Henry Taube is one of the most creative exchange reaction between Pb2+ and Pb4+ (7).