Ted @5350” Rams
Total Page:16
File Type:pdf, Size:1020Kb

Load more
Recommended publications
-

Indoor Plants Or Houseplants
Visit us on the Web: www.gardeninghelp.org Indoor Plants or Houseplants Over the past twenty years houseplants have grown in popularity. Offered in a wide variety of sizes, shapes, colors and textures, houseplants beautify our homes and help soften our environment. They have been scientifically proven to improve our health by lowering blood pressure and removing pollutants from the air we breathe. When selecting a houseplant, choose reputable suppliers who specialize in growing houseplants. Get off to a good start by thoroughly examining each plant. Watch for brown edges and spindly growth with elongated stems and large gaps between new leaves. Inspect leaves and stem junctions for signs of insect or disease problems. Check any support stakes to make sure they are not hiding broken stems or branches. Finally, make sure the plant is placed in an area that suits its optimal requirements for light, temperature and humidity. Where to Place Your House Plants With the exception of the very darkest areas, you can always find a houseplant with growth requirements to match the environmental conditions in your home. The most important factors are light intensity and duration. The best way to determine the intensity of light at a window exposure area is to measure it with a light meter. A light meter measures light in units called foot-candles. One foot-candle is the amount of light from a candle spread over a square foot of surface area. Plants that prefer low light may produce dull, lifeless-looking leaves when exposed to bright light. Bright light can also cause leaf spots or brown-tipped scorched margins. -

Buy Pilea Cadierei, Pilea Aluminium, Watermelon Pilea - Plant Online at Nurserylive | Best Plants at Lowest Price
Buy pilea cadierei, pilea aluminium, watermelon pilea - plant online at nurserylive | Best plants at lowest price Pilea Cadierei, Pilea Aluminium, Watermelon Pilea - Plant Showy, silver-splashed leaves make Aluminum Plant a stunning and popular house plant. Rating: Not Rated Yet Price Variant price modifier: Base price with tax Price with discount ?299 Salesprice with discount Sales price ?299 Sales price without tax ?299 Discount Tax amount Ask a question about this product Description With this purchase you will get: 01 Pilea Cadierei, Pilea Aluminium, Watermelon Pilea Plant 01 6 inch Grower Round Plastic Pot (Black) Description for Pilea Cadierei, Pilea Aluminium, Watermelon Pilea Plant height: 9 - 15 inches (22 - 39 cm) 1 / 3 Buy pilea cadierei, pilea aluminium, watermelon pilea - plant online at nurserylive | Best plants at lowest price Plant spread: Pilea cadierei is a species of flowering plant in the family Urticaceae, native to China and Vietnam. It is an evergreen perennial growing up to 30 cm tall by 21 cm broad, with dark green oval leaves, each leaf having four raised silvery patches. Common name(s): Aluminium Plant, Watermelon Plant Flower colours: Green, white Bloom time: Late Fall, Early Winter Max reachable height: 0.75 to 1.00 feet Difficulty to grow: Easy to grow Planting and care Common is a vine and has larger glossy green leaves than Royal. Both can survive in temperate climates if they are planted in a sheltered area. Arabian is a small bush with evergreen leaves. There are many other varieties of plant, of which are best suited for sub-tropical climates. -

Pilea Cadierei Aluminum Plant1 Edward F
FPS478 Pilea cadierei Aluminum Plant1 Edward F. Gilman2 Introduction Uses: hanging basket; suitable for growing indoors; ground cover; cascading down a wall The variegated foliage on aluminum plant is unlike any Availability: generally available in many areas within its other, with shiny silver, irregularly shaped markings parallel hardiness range to the lateral veins (Fig. 1). Leaves are held opposite each other on square, green stems, producing a thick ground cover about 12 inches tall in a shaded landscape. Small, white flowers are produced at the ends of the stems in the summer, but they are mostly overshadowed by the conspicuous foliage. Figure 2. Shaded area represents potential planting range. Description Figure 1. Aluminum plant Height: .5 to 1 feet General Information Spread: depends upon supporting structure Plant habit: spreading Scientific name: Pilea cadierei Plant density: moderate Pronunciation: PYE-lee-uh kuh-DEER-ree-eye Growth rate: moderate Common name(s): aluminum plant Texture: medium Family: Urticaceae Plant type: ground cover Foliage USDA hardiness zones: 10 through 11 (Fig. 2) Planting month for zone 10 and 11: year round Leaf arrangement: opposite/subopposite Origin: not native to North America Leaf type: simple Leaf margin: serrate 1. This document is FPS478, one of a series of the Environmental Horticulture Department, UF/IFAS Extension. Original publication date October 1999. Reviewed February 2014. Visit the EDIS website at http://edis.ifas.ufl.edu. 2. Edward F. Gilman, professor, Environmental Horticulture Department; UF/IFAS Extension, Gainesville, FL 32611. The Institute of Food and Agricultural Sciences (IFAS) is an Equal Opportunity Institution authorized to provide research, educational information and other services only to individuals and institutions that function with non-discrimination with respect to race, creed, color, religion, age, disability, sex, sexual orientation, marital status, national origin, political opinions or affiliations. -

Brief Note: Preliminary Investigation of Air Blisters in Pilea Cadierei
Copyright © 1980 Ohio Acad. Sci. 0030-0950/80/0006-0280 $1.00/0 BRIEF NOTE PRELIMINARY INVESTIGATION OF AIR BLISTERS IN PILEA CADIEREI1 BRIAN D. DOWNS, KEVIN C. VAUGHN and KENNETH G. WILSON, Dept. of Botany, Miami Univ., Oxford, OH 45056 OHIO J. SCI. 80(6): 280, 1980 A form of highly ornamental variega- amined with an Hitachi HS-9 electron tion in higher plants known as "air microscope. blisters" (Kirk and Tilney-Bassett 1967) Mueller and Greenwood (1978) ob- in which the leaves have silver or cream- served that phenolic-storing cells con- colored dotting or striping patterns is tained a blackened, osmiophilic cyto- very common. Species of the genera plasm due to the leaching of the phenolics Pilea, Begonia, Episcia, Dioscorea, Ze- from the vacuoles, obscuring cellular de- brina and Trifolium, to name but a few, tail and making any ultrastructural study have distinct air blisters. As common difficult. This same blackening was as this condition appears to be, it is per- found in the subepidermal air blister and haps surprising that it has not been ex- thus possibly indicated that these cells tensively studied. Hara (1957) investi- are also phenolic-storing cells. Figure 1 gated these plants by light microscopy shows a dark, osmiophilic phenolic-stor- but did no physiological or subcellular ing cell of the air blister of Pilea cadierei studies, and genetic data regarding these with relatively electron transparent epi- plants were summarized by Kirk and dermal and other cells. The plastids and Tilney-Bassett (1967). No likely role other organelles contained in the air for these air blisters has been considered blister cells are highly osmiophylic due to in either work. -

Updating Material of Study Subject Flower
LANDSCAPING FACULTY DEPARTMENT OF GARDENING AND AGRICULTURAL TECHNOLOGIES STUDY PROGRAMME: GARDENING TERRITORIES AND THEIR DESIGN (code) 653H93002 LAIMA MARKEVIČIENĖ UPDATING MATERIAL OF STUDY SUBJECT FLOWER GROWING (IN GREENHOUSES) ATNAUJINIMO MEDŽIAGA PROJEKTUI TO SUPPORT THE PROJECT ‘INTERNATIONAL DEVELOPMENT IN THE ENGINEERING STUDY FIELD PROGRAMMES AND THEIR UPDATING BY CUSTOMIZING TO MEET COURSE DELIVERY NEEDS OF INTERNATIONAL STUDENTS AT THE LANDSCAPING FACULTY OF KAUNO KOLEGIJA/UNIVERSITY OF APPLIED SCIENCES‘ (VP1-2.2-ŠMM-07-K-02-045) Mastaičiai 2012 Educational Institution: Kauno Kolegija / University of Applied Sciences Study Programme: Growing Territories and their Design Study Subject Programme FLOWER GROWING 1. The Annotation. Study Field Subject, in which decorative, morphological and bioecological characteristics of annual, biennial, perennial, bulbous, room: greenhouses and interior flowers are analyzed. Knowledge and abilities when evaluating and applying them in growing territories and interior are given. 2. The Aim of the Programme. To describe and evalaute grass decorative plants, by choosing them for growing territories and interiors of different types, to develope the skills of plants researches and holistic attitude when performing professional solutions. 3.The Length in Credits and Hours: Structure Length Practical Study in Lectures, Consultations, Individual In total: works, Assessment subject ECTS hours hours work, hours hours hours title credits Flower growing 12 69 72 19 160 320 1. Outside 6 29 39 12 80 160 Examination 2. Room 6 40 33 7 80 160 Examination 4.Prerequisites: Chemistry and Plants Protection, Fundamentals of Agronomy, Information Technologies, Foreign Language. 5. Links between Learning Outcomes and Intended Study Subject Outcomes and Student Achievement Assessment Methods: Learning outcomes Intended study subject Student achievement Study methods outcomes assessment methods Lecture, telling, explanation, Testing, frontal inquiry, 1. -

A-1-17 Plants for Terrariums and Dish Gardens.Pmd
HOME GROUNDS FACT SHEET Horticulture Center Cornell University Demonstration & Community Gardens at East Meadow Farm Cooperative Extension 832 Merrick Avenue East Meadow, NY 11554 Nassau County Phone: 516-565-5265 Plants for Terrariums and Dish Gardens Invented in 1836 by N. B. Ward and originally Soil called “Wardian Cases,” terrariums were used for Soil from the garden is not good for terrariums. A transporting growing plants on long sea voyages. good soil mix should be composed of 1/3 soil, 1/3 They provided the best and safest method of sand and 1/3 peat moss. Some houseplant mixes transporting potted and living plants across the sold under various trade names are ideal. One ocean. tablespoon of water soluble houseplant food in dry A true terrarium is an enclosed environment. form may be added to 2 pounds or 1 quart of soil When it is properly sited so it receives the correct mix. When you add soil to the terrarium, make sure light, it should be almost self-sustaining for at least it's no more than 1/5 the total volume of the ter- a month, perhaps longer. rarium. Since there are no drainage holes, some The terrarium can be a work of art when the provision must be made for this. A layer of sand, proper materials are cleverly combined in an small gravel or moss can be placed on the bottom attractive container. It can also provide the for drainage. opportunity to grow some of the delicate plants that would ordinarily dwindle rapidly when exposed to the dry atmosphere of the home. -
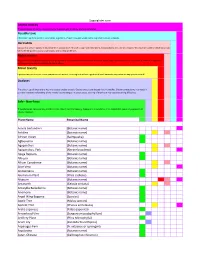
Plant Name Botanical Name Acacia Berlandieri
Sugarglider.com Animal toxicity These plants are known to cause problems in animals and should be avoided completely. Possibly toxic Information on these plants is incomplete. Ingestions of small amounts would not be expected to cause problems. Dermatitis Exposure or a thorn puncture wound when in contact with the juice or sap from these plants may produce a skin rash or irritation. Wounds from some of these plants can be extremely painful causing large blisters and burning of the skin. Major toxicity Ingestions of these plants, especially in large amounts, are expected to cause serious effects to major body organs such as the liver, heart or kidneys. If ingested, call the poison center immediately for more advice. Minor toxicity Ingestion may cause some minor symptoms such as rash, vomiting or diarrhea. Ingestion of small amounts may not cause any symptoms at all. Oxalates The juice or sap of these plants has microscopic oxalate crystals. Oxalate crystals are shaped like tiny needles. Chewing these plants may result in pain and irritation and swelling of the mouth, lips and tongue. In severe cases, swelling of the throat may cause breathing difficulties. Safe - Non-Toxic These plants are not poisonous and there is no known record of toxicity. Exposure to these plants is not expected to cause any symptoms or adverse reactions. Plant Name Botanical Name Acacia berlandieri (Botanic name) Achillea (Botanic name) African Violet (Saintpaulia) Aglaonema (Botanic name) Agapanthus (Botanic name) Agapanthus, Pink (Nerine bowdenii) Ajuga Reptans (Botanic name) Albizzia (Botanic name) Allium Canadense (Botanic name) Aloe Vera (Botanic name) Alstoemeria (Botanic name) Aluminum Plant (Pilea cadierei) Alyssum (Botanic name) Amaranth (Celosia cristata) Amaryllis Belladonna (Botanic name) Anemone (Botanic name) Angel Wing Begonia (Species) Apple Tree (Malus species) Apricot Tree (Prunus armeniaca) Aralia Japanese (Fatsia japonica) Arrowhead Vine (Syngonium podophyllum) Artillery Plant (Pilea Microphylla) Arum Lily (Zantedschia lethipica) Asparagus Fern (A. -

Pilea Cadierei1
Fact Sheet FPS-478 October, 1999 Pilea cadierei1 Edward F. Gilman2 Introduction The variegated foliage on Aluminum Plant is unlike any other, with shiny silver, irregularly-shaped markings parallel to the lateral veins (Fig. 1). Leaves are held opposite each other on square, green stems producing a thick ground cover about 12-inches-tall in a shaded landscape. Small white flowers are produced at the ends of the stems in the summer but they are mostly overshadowed by the conspicuous foliage. General Information Scientific name: Pilea cadierei Pronunciation: PYE-lee-uh kuh-DEER-ree-eye Common name(s): Aluminum Plant Family: Urticaceae Plant type: ground cover USDA hardiness zones: 10 through 11 (Fig. 2) Planting month for zone 10 and 11: year round Origin: not native to North America Uses: hanging basket; suitable for growing indoors; ground cover; cascading down a wall Figure 1. Aluminum Plant. Availablity: generally available in many areas within its hardiness range Texture: medium Description Foliage Height: .5 to 1 feet Spread: depends upon supporting structure Leaf arrangement: opposite/subopposite Plant habit: spreading Leaf type: simple Plant density: moderate Leaf margin: serrate Growth rate: moderate Leaf shape: obovate 1.This document is Fact Sheet FPS-478, one of a series of the Environmental Horticulture Department, Florida Cooperative Extension Service, Institute of Food and Agricultural Sciences, University of Florida. Publication date: October, 1999 Please visit the EDIS Web site at http:/edis.ifas.ufl.edu. 2. Edward F. Gilman, professor, Environmental Horticulture Department, Cooperative Extension Service, Institute of Food and Agricultural Sciences, University of Florida, Gainesville, 32611. -

Molecular Phylogenetics and Character Evolution of Cannabaceae
TAXON 62 (3) • June 2013: 473–485 Yang & al. • Phylogenetics and character evolution of Cannabaceae Molecular phylogenetics and character evolution of Cannabaceae Mei-Qing Yang,1,2,3 Robin van Velzen,4,5 Freek T. Bakker,4 Ali Sattarian,6 De-Zhu Li1,2 & Ting-Shuang Yi1,2 1 Key Laboratory of Biodiversity and Biogeography, Kunming Institute of Botany, Chinese Academy of Sciences, Kunming, Yunnan 650201, P.R. China 2 Plant Germplasm and Genomics Center, Germplasm Bank of Wild Species, Kunming Institute of Botany, Chinese Academy of Sciences, Kunming, Yunnan 650201, P.R. China 3 University of Chinese Academy of Sciences, Beijing 100093, P.R. China 4 Biosystematics Group, Wageningen University, Droevendaalsesteeg 1, 6708 PB Wageningen, The Netherlands 5 Laboratory of Molecular Biology, Wageningen University, Droevendaalsesteeg 1, 6708 PB Wageningen, The Netherlands 6 Department of Natural Resources, Gonbad University, Gonbad Kavous 4971799151, Iran Authors for correspondence: Ting-Shuang Yi, [email protected]; De-Zhu Li, [email protected] Abstract Cannabaceae includes ten genera that are widely distributed in tropical to temperate regions of the world. Because of limited taxon and character sampling in previous studies, intergeneric phylogenetic relationships within this family have been poorly resolved. We conducted a molecular phylogenetic study based on four plastid loci (atpB-rbcL, rbcL, rps16, trnL-trnF) from 36 ingroup taxa, representing all ten recognized Cannabaceae genera, and six related taxa as outgroups. The molecular results strongly supported this expanded family to be a monophyletic group. All genera were monophyletic except for Trema, which was paraphyletic with respect to Parasponia. The Aphananthe clade was sister to all other Cannabaceae, and the other genera formed a strongly supported clade further resolved into a Lozanella clade, a Gironniera clade, and a trichotomy formed by the remaining genera. -

Pilea Cadierei F
Pilea cadierei F. Gagnepain & A. Guillaumin Aluminum Plant Other Common Names: Watermelon Pilea. Family: Urticaceae. Cold Hardiness: Plants are grown outdoors in USDA plant hardiness zones 10 (9) to 11. Foliage: Evergreen, opposite, simple, 2O to 3O long elliptic to obovate leaves have nearly entire to slightly scalloped or crenate margins, rounded to obtuse bases, and acute to acuminate tips; the glossy blades are mottled white, silver (aluminum colored), and green above; the underside of new leaves are rusty red as they initially unfold; venation is palmate with three main veins running the length of the leaf; tiny whitish linear calcium car- bonate secretions (cystoliths) form on the leaves; veins are green with mottling appearing principally between the veins; short stout petioles are yellow-green, sometimes with a reddish flush, and flattened to slightly con- cave above. Flower: The small loose axillary cymes of very small individual flowers are not ornamentally important. Fruit: The tiny achenes are not ornamental. Stem / Bark: Stems — succulent dark green stems with brown lenticels have pairs of leaves perpendicular to the next pair above and below; Buds — foliose, red-green; Bark — not applicable. Habit: Aluminum Plant is an erect herbaceous perennial which can quickly grow to a height of 12O to 18O, howev- er the most common selections in cultivation mature at a shorter height of half this size; it quickly forms a groundcover by rooting where branches touch the soil; the overall texture is medium to medium-coarse. Cultural Requirements: Aluminum Plant can be easily grown in warm climates, or as a seasonal annual, in most locations in which moderate shade, moist well drained soils, and sufficient high quality irrigation water are available; better fertility increases foliage growth which is the whole point of growing this species; a location with protection from winter freeze and wind will improve chances of plants returning from the roots out- doors in marginal USDA zone 9 locations. -

PEST ALERT Pest Alert Created-November-2018 Florida Department of Agriculture and Consumer Services Division of Plant Industry
FDACS-P-01916 PEST ALERT Pest Alert created-November-2018 Florida Department of Agriculture and Consumer Services Division of Plant Industry First Report of the Pilea Aphid (Myzus fataunae Shinji) in the Western Hemisphere Katherine E. O. Fairbanks, Susan E. Halbert; Bureau of Entomology, Nematology and Plant Pathology [email protected] or 1-888-397-1517 INTRODUCTION: On August 22, 2018, from Longwood, Seminole County, Florida, Jesse Krok (DPI) collected a sample of aluminum plant (Pilea cadeirei Gagnep. & Guill.) infested with all life stages of the aphid Myzus fataunae Shinji 1924 (after Takahashi 1965). Myzus fataunae is native to Japan and Korea; this represents the first record of the species in the Western Hemisphere. HOSTS: The species was described originally from Fatoua (Moraceae) (Shinji 1924), though subsequent collections have been from members of the Urticaceae: aluminum plant, Pilea cadierei Gagnep. & Guill.; Canadian or Japanese clearweed, Pilea pumila (hamaoi) (L.) A. Gray; and other ornamental Pilea, Boehmeria and Parietaria species (Takahashi 1965, Miyazaki 1971, Blackman & Eastop 2018). SURVEY: A hand lens is necessary to survey for M. fataunae. Other aphids may be present on the host and potential host plants, but the target will stand out by its small size and distinct bicoloration. Myzus fataunae appears to feed on all above- ground parts of the plant, but favors new growth and petioles. Cut potentially infested plant parts and place them in a vial of alcohol. Alternatively, separate foliage from roots (to prevent soil particles from obscuring aphids) and beat it against the side of a shallow white tray; then, tap the plant material over the tray using a pencil. -

Downloaded from Genbank (Table1) Were Aligned with the Online Version of MAFFT V
Journal of Fungi Article Molecular and Morphological Assessment of Septoria Species Associated with Ornamental Plants in Yunnan Province, China Yuan-Yan An 1, Monika C. Dayarathne 1, Xiang-Yu Zeng 1,2, Alan J. L. Phillips 3 , Kevin D. Hyde 2,4 and Yong Wang 1,* 1 Department of Plant Pathology, College of Agriculture, Guizhou University, Guiyang 550025, China; [email protected] (Y.-Y.A.); [email protected] (M.C.D.); [email protected] (X.-Y.Z.) 2 Center of Excellence in Fungal Research, Mae Fah Luang University, Chiang Rai 57100, Thailand; [email protected] 3 Faculdade de Ciências, Biosystems and Integrative Sciences Institute (BioISI), Universidade de Lisboa, Campo Grande, 1749-016 Lisbon, Portugal; [email protected] 4 Institute of Plant Health, Zhongkai, University of Agriculture and Engineering, Haizhu District, Guangzhou 510225, China * Correspondence: [email protected]; Tel.: +86-0187-9889-9302 Abstract: The Karst landform is the main geographic characteristic in South China. Such areas are rich in vegetation and especially suitable for growth of shrubs and herbaceous plants. In this study, 11 Septoria strains were obtained from different plants’ leaves collected in the Kunming Botanical Garden, Yunnan Province, China. Based on single-gene and multi-gene analyses of five gene loci (tef1, rpb2, tub2, ITS, and LSU) and four gene regions (without LSU), these strains were found to belong to three independent phylogenetic lineages representing five species, including four novel taxa, and one new record for China. Five single gene trees were also provided to evaluate the Citation: An, Y.-Y.; Dayarathne, effectiveness of each gene for discriminating the species, as a result of which tub2 was found to have M.C.; Zeng, X.-Y.; Phillips, A.J.L.; the most suitable DNA barcode for rapid identification.