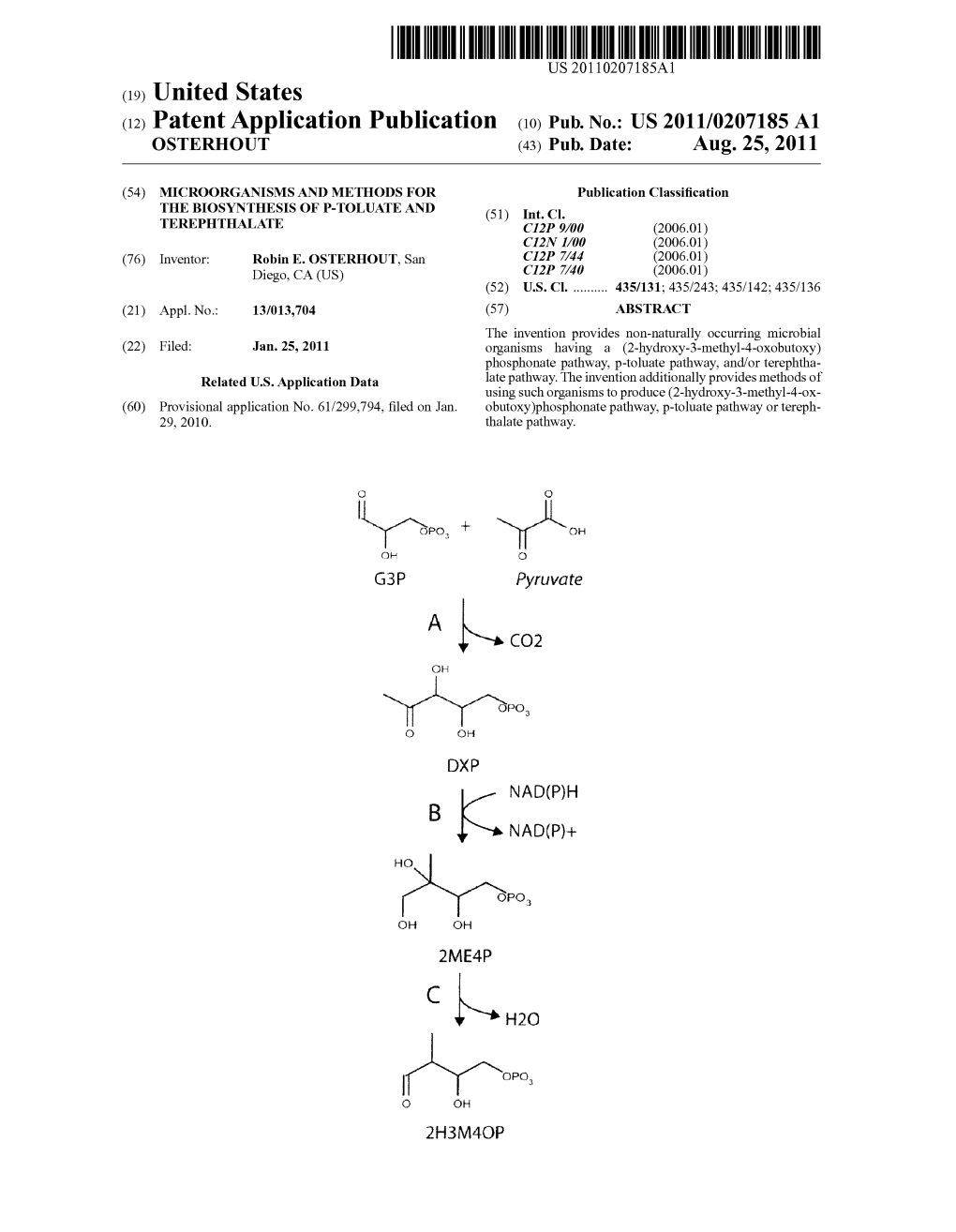
Sirso,O O O DXP NAD(P)H B N AD(P)+
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

De Novo Sequencing and Transcriptome Analysis Reveal Key Genes Regulating Steroid Metabolism in Leaves, Roots, Adventitious Roots and Calli of Periploca Sepium Bunge
ORIGINAL RESEARCH published: 21 April 2017 doi: 10.3389/fpls.2017.00594 De novo Sequencing and Transcriptome Analysis Reveal Key Genes Regulating Steroid Metabolism in Leaves, Roots, Adventitious Roots and Calli of Periploca sepium Bunge Jian Zhang 1, 2, 3, Xinglin Li 1, 3*, Fuping Lu 1, 3, Shanying Wang 1, 3, Yunhe An 4, Xiaoxing Su 4, Xiankuan Li 2, Lin Ma 2 and Guangjian Han 5 1 Key Lab of Industrial Fermentation Microbiology, Tianjin University of Science and Technology, Ministry of Education, Tianjin, China, 2 School of Traditional Chinese Materia Medica, Tianjin University of Traditional Chinese Medicine, Tianjin, China, 3 College of Bioengineering, Tianjin University of Science and Technology, Tianjin, China, 4 Beijing Center for Physical and Chemical Analysis, Beijing, China, 5 Shachuan Biotechnology, Tianjin, China Edited by: Periploca sepium Bunge is a traditional medicinal plant, whose root bark is important Peng Zhang, Institute of Plant Physiology and for Chinese herbal medicine. Its major bioactive compounds are C21 steroids and Ecology, SIBS, CAS, China periplocin, a kind of cardiac glycoside, which are derived from the steroid synthesis Reviewed by: pathway. However, research on P. sepium genome or transcriptomes and their related Kun Yu, genes has been lacking for a long time. In this study we estimated this species Hubei University of Chinese Medicine, China nuclear genome size at 170 Mb (using flow cytometry). Then, RNA sequencing of Jun Yang, four different tissue samples of P. sepium (leaves, roots, adventitious roots, and Shanghai Chenshan Plant Science Research Center (CAS), China calli) was done using the sequencing platform Illumina/Solexa Hiseq 2,500. -

Structural and Biochemical Characterizations of Three Potential Drug Targets from Pathogens
Digital Comprehensive Summaries of Uppsala Dissertations from the Faculty of Science and Technology 2020 Structural and Biochemical Characterizations of Three Potential Drug Targets from Pathogens LU LU ACTA UNIVERSITATIS UPSALIENSIS ISSN 1651-6214 ISBN 978-91-513-1148-7 UPPSALA urn:nbn:se:uu:diva-435815 2021 Dissertation presented at Uppsala University to be publicly examined in Room A1:111a, BMC, Husargatan 3, Uppsala, Friday, 16 April 2021 at 13:15 for the degree of Doctor of Philosophy. The examination will be conducted in English. Faculty examiner: Christian Cambillau. Abstract Lu, L. 2021. Structural and Biochemical Characterizations of Three Potential Drug Targets from Pathogens. Digital Comprehensive Summaries of Uppsala Dissertations from the Faculty of Science and Technology 2020. 91 pp. Uppsala: Acta Universitatis Upsaliensis. ISBN 978-91-513-1148-7. As antibiotic resistance of various pathogens emerged globally, the need for new effective drugs with novel modes of action became urgent. In this thesis, we focus on infectious diseases, e.g. tuberculosis, malaria, and nosocomial infections, and the corresponding causative pathogens, Mycobacterium tuberculosis, Plasmodium falciparum, and the Gram-negative ESKAPE pathogens that underlie so many healthcare-acquired diseases. Following the same- target-other-pathogen (STOP) strategy, we attempted to comprehensively explore the properties of three promising drug targets. Signal peptidase I (SPase I), existing both in Gram-negative and Gram-positive bacteria, as well as in parasites, is vital for cell viability, due to its critical role in signal peptide cleavage, thus, protein maturation, and secreted protein transport. Three factors, comprising essentiality, a unique mode of action, and easy accessibility, make it an attractive drug target. -

Sauret-Gü Eto S
Mutations in Escherichia coli aceE and ribB Genes Allow Survival of Strains Defective in the First Step of the Isoprenoid Biosynthesis Pathway Jordi Perez-Gil1, Eva Maria Uros1, Susanna Sauret-Gu¨ eto1¤, L. Maria Lois1, James Kirby2, Minobu Nishimoto2, Edward E. K. Baidoo2, Jay D. Keasling2, Albert Boronat1,3, Manuel Rodriguez- Concepcion1* 1 Centre for Research in Agricultural Genomics (CRAG) CSIC-IRTA-UAB-UB, Campus UAB Bellaterra, Barcelona, Spain, 2 Joint BioEnergy Institute, Emeryville, California, United States of America, 3 Department de Bioquı´mica i Biologia Molecular, Universitat de Barcelona, Barcelona, Spain Abstract A functional 2-C-methyl-D-erythritol 4-phosphate (MEP) pathway is required for isoprenoid biosynthesis and hence survival in Escherichia coli and most other bacteria. In the first two steps of the pathway, MEP is produced from the central metabolic intermediates pyruvate and glyceraldehyde 3-phosphate via 1-deoxy-D-xylulose 5-phosphate (DXP) by the activity of the enzymes DXP synthase (DXS) and DXP reductoisomerase (DXR). Because the MEP pathway is absent from humans, it was proposed as a promising new target to develop new antibiotics. However, the lethal phenotype caused by the deletion of DXS or DXR was found to be suppressed with a relatively high efficiency by unidentified mutations. Here we report that several mutations in the unrelated genes aceE and ribB rescue growth of DXS-defective mutants because the encoded enzymes allowed the production of sufficient DXP in vivo. Together, this work unveils the diversity of mechanisms that can evolve in bacteria to circumvent a blockage of the first step of the MEP pathway. -

The Coordinated Upregulated Expression of Genes Involved In
plants Article The Coordinated Upregulated Expression of Genes Involved in MEP, Chlorophyll, Carotenoid and Tocopherol Pathways, Mirrored the Corresponding Metabolite Contents in Rice Leaves during De-Etiolation Xin Jin 1,2,3,†, Can Baysal 3,†, Margit Drapal 4, Yanmin Sheng 5, Xin Huang 3, Wenshu He 3, Lianxuan Shi 6, Teresa Capell 3, Paul D. Fraser 4, Paul Christou 3,7 and Changfu Zhu 3,5,* 1 Gansu Provincial Key Laboratory of Aridland Crop Science, Gansu Agricultural University, Lanzhou 730070, China; [email protected] 2 College of Life Science and Technology, Gansu Agricultural University, Lanzhou 730070, China 3 Department of Plant Production and Forestry Science, University of Lleida-Agrotecnio CERCA Center, Av. Alcalde Rovira Roure, 191, 25198 Lleida, Spain; [email protected] (C.B.); [email protected] (X.H.); [email protected] (W.H.); [email protected] (T.C.); [email protected] (P.C.) 4 Biochemistry, School of Life Sciences and Environment, Royal Holloway University of London, Egham Hill, Egham, Surrey TW20 0EX, UK; [email protected] (M.D.); [email protected] (P.D.F.) 5 School of Life Sciences, Changchun Normal University, Changchun 130032, China; [email protected] 6 School of Life Sciences, Northeast Normal University, Changchun 130024, China; [email protected] 7 ICREA, Catalan Institute for Research and Advanced Studies, Passeig Lluís Companys 23, 08010 Barcelona, Spain Citation: Jin, X.; Baysal, C.; Drapal, * Correspondence: [email protected] M.; Sheng, Y.; Huang, X.; He, W.; Shi, † These authors contributed equally to this work. L.; Capell, T.; Fraser, P.D.; Christou, P.; et al. -

Evolutionary Diversification and Characterization of the Eubacterial
Carretero-Paulet et al. BMC Evolutionary Biology 2013, 13:180 http://www.biomedcentral.com/1471-2148/13/180 RESEARCH ARTICLE Open Access Evolutionary diversification and characterization of the eubacterial gene family encoding DXR type II, an alternative isoprenoid biosynthetic enzyme Lorenzo Carretero-Paulet1,2*, Agnieszka Lipska1, Jordi Pérez-Gil3, Félix J Sangari4, Victor A Albert1 and Manuel Rodríguez-Concepción3* Abstract Background: Isoprenoids constitute a vast family of natural compounds performing diverse and essential functions in all domains of life. In most eubacteria, isoprenoids are synthesized through the methylerythritol 4-phosphate (MEP) pathway. The production of MEP is usually catalyzed by deoxyxylulose 5-phosphate reductoisomerase (DXR-I) but a few organisms use an alternative DXR-like enzyme (DXR-II). Results: Searches through 1498 bacterial complete proteomes detected 130 sequences with similarity to DXR-II. Phylogenetic analysis identified three well-resolved clades: the DXR-II family (clustering 53 sequences including eleven experimentally verified as functional enzymes able to produce MEP), and two previously uncharacterized NAD(P)-dependent oxidoreductase families (designated DLO1 and DLO2 for DXR-II-like oxidoreductases 1 and 2). Our analyses identified amino acid changes critical for the acquisition of DXR-II biochemical function through type-I functional divergence, two of them mapping onto key residues for DXR-II activity. DXR-II showed a markedly discontinuous distribution, which was verified at several levels: taxonomic (being predominantly found in Alphaproteobacteria and Firmicutes), metabolic (being mostly found in bacteria with complete functional MEP pathways with or without DXR-I), and phenotypic (as no biological/phenotypic property was found to be preferentially distributed among DXR-II-containing strains, apart from pathogenicity in animals). -

Supporting Information for Proteomics DOI 10.1002/Pmic.200600568
Supporting Information for Proteomics DOI 10.1002/pmic.200600568 Xiao-Li Wang, Alex Fu, Sreekumar Raghavakaimal and Hon-Chi Lee Proteomic analysis of vascular endothelial cells in response to laminar shear stress ª 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com Table A. Proteins altereations after 10 minutes of shear stress Accesion # Protein Name H:L* Matched Sequence Charge Score Confidence % Error Peptide Number gi|39931056 1-deoxy-D-xylulose 5-phosphate reductoisomerase 1 (DXP reductoisomerase 1) 3.78 QNGCR 2 21 75 1.6 2 gi|14548081 28 kDa and 33 kDa Inner dynein arm light chain, axonemal (p28, p33) 2.86 CEAIEK 2 32 90 3.7 3 gi|1723793 3 beta-hydroxysteroid dehydrogenase 0.21 VKIVCAYR 2 28 90 5.9 2 gi|32171609 30S ribosomal protein S11 2.17 GCAHIK 2 33 90 6.2 4 gi|34222742 30S ribosomal protein S2 0.41 REYNAVAECHK 3 20 75 1.3 4 gi|26399495 30S ribosomal protein S5P 0.19 GCGSWEZGZGRPHTVALK 3 25 75 1 4 gi|3929352 3-hydroxy-3-methylglutaryl-coenzyme A reductase (HMG-CoA reductase) 0.07 GCSVITR 2 23 75 3.2 4 gi|23396668 3-isopropylmalate dehydratase small subunit (Isopropylmalate isomerase) 0.46 AAGVSCVIAK 2 25 75 1.3 3 gi|3123178 50S ribosomal protein L32 (Ribosomal protein I) (BL37) 2.17 ACGTYK 2 32 90 6.2 2 gi|135758 Acetyl-CoA acetyltransferase (Acetoacetyl-CoA thiolase) 0.31 VCASAMK 2 20 75 1.2 3 gi|14194435 ADAM 11 precursor (A disintegrin and metalloproteinase domain 11) 0.23 GSCGRK 2 24 75 4 2 gi|6174971 Adenosylhomocysteinase (S-adenosyl-L-homocysteine hydrolase) (AdoHcyase) 2.86 GCAQALK 2 32 90 3.7 -

Vitamin B6 Biosynthesis in Higher Plants
Vitamin B6 biosynthesis in higher plants Marina Tambasco-Studart*, Olca Titiz*, Thomas Raschle, Gabriela Forster, Nikolaus Amrhein, and Teresa B. Fitzpatrick† Institute of Plant Sciences, Swiss Federal Institute of Technology (ETH), Universita¨tstrasse 2, CH-8092 Zu¨rich, Switzerland Communicated by Meinhart H. Zenk, Universita¨t Halle, Halle͞Saale, Germany, July 21, 2005 (received for review May 31, 2005) Vitamin B6 is an essential metabolite in all organisms. It can act as a that does not involve DXP exists for vitamin B6 biosynthesis. Only coenzyme for numerous metabolic enzymes and has recently been two genes (PDX1 and PDX2), which show no homology to any of shown to be a potent antioxidant. Plants and microorganisms have the E. coli genes, appear to be involved in this alternative pathway a de novo biosynthetic pathway for vitamin B6, but animals must (2, 15). Furthermore, based on extensive genomic analyses, it has obtain it from dietary sources. In Escherichia coli, it is known that been shown that the pathways of vitamin B6 biosynthesis are the vitamin is derived from deoxyxylulose 5-phosphate (an inter- autoexclusive in that organisms have the genes for one or the other mediate in the nonmevalonate pathway of isoprenoid biosynthe- pathway but not both (2, 15). PDX1 and PDX2 have been predicted sis) and 4-phosphohydroxy-L-threonine. It has been assumed that to function as a glutamine amidotransferase with PDX2 as the vitamin B6 is synthesized in the same way in plants, but this glutaminase domain and PDX1 as the acceptor͞synthase domain. hypothesis has never been experimentally proven. Here, we show Indeed, glutaminase activity has been demonstrated for PDX2 that, in plants, synthesis of the vitamin takes an entirely different from a number of organisms (16–18). -

A 1-Deoxy-D-Xylulose 5-Phosphate Reductoisomerase Catalyzing The
Proc. Natl. Acad. Sci. USA Vol. 95, pp. 9879–9884, August 1998 Biochemistry A 1-deoxy-D-xylulose 5-phosphate reductoisomerase catalyzing the formation of 2-C-methyl-D-erythritol 4-phosphate in an alternative nonmevalonate pathway for terpenoid biosynthesis SHUNJI TAKAHASHI*, TOMOHISA KUZUYAMA*, HIROYUKI WATANABE, AND HARUO SETO† Institute of Molecular and Cellular Biosciences, University of Tokyo, Bunkyo-ku, Tokyo 113-0032, Japan Communicated by Christopher T. Walsh, Harvard Medical School, Boston, MA, June 22, 1998 (received for review May 15, 1998) ABSTRACT Several eubacteria including Esherichia coli 3-phosphate (Fig. 1A). In the second step the intramolecular use an alternative nonmevalonate pathway for the biosynthe- rearrangement of DXP is assumed to give a hypothetical sis of isopentenyl diphosphate instead of the ubiquitous rearrangement intermediate 2-C-methylerythrose 4-phos- mevalonate pathway. In the alternative pathway, 2-C-methyl- phate, which is then converted to 2-C-methyl-D-erythritol D-erythritol or its 4-phosphate, which is proposed to be 4-phosphate (MEP) by an unspecified reduction process (7). formed from 1-deoxy-D-xylulose 5-phosphate via intramolec- Thus, precise details about the reductive mechanism for the ular rearrangement followed by reduction process, is one of formation of MEP remained to be unclear. the biosynthetic precursors of isopentenyl diphosphate. To A precedent for this biosynthetic rearrangement reaction is clone the gene(s) responsible for synthesis of 2-C-methyl-D- found in the biosynthesis of valine, isoleucine, and leucine. The erythritol 4-phosphate, we prepared and selected E. coli enzyme involved in this reaction ketol acid reductoisomerase mutants with an obligatory requirement for 2-C-methyleryth- (EC1.1.1.86) catalyzes rearrangement of 2-acetolactate to ritol for growth and survival. -

Oral Metapangenomics Daniel Utter 13 January 2020
Oral Metapangenomics Daniel Utter 13 January 2020 Contents Overview . .1 Organization & Workflow . .2 Step 1 - Collecting and annotating genomes . .2 Getting the genomes from NCBI . .2 Anvi’o contigs database . .3 Whole thing . .3 Functional annotation . .4 Step 2 - Mapping . .5 Data acquisition - HMP Metagenomes . .5 Recruiting HMP metagenomes to the contigs . .6 A short statement on competitive mapping and its interpretation . .7 Step 3 - Profiling the metagenome recruitment . .8 Step 4 - Pangenome and metapangenome construction . .9 Some background on the methods wrapped by the anvio programs . 10 Combining metapangenomes . 11 Anvi’o interactive display choices for metapangenomes . 13 Supplemental tables . 13 Querying the gene cluster annotations for enriched functions . 13 Granular analysis of mapping results at the per-gene, per-sample level . 17 Differential function analysis . 20 Nucleotide level coverage plots (Figure 4A, 4B) . 21 Rothia candidate driver of adaptation to BM . 21 Investigating the effect of non-specific mapping . 25 Overview This is the long-form, narrative version of the metapangenomic methods used for our study Metapange- nomics. Our goal in writing this is to make the workflow be as transparent and reducible as possible, and also to provide a step-by-step workflow for interested readers to adapt our methods to their own needs. Most of the code here is based off of Tom Delmont and Meren’s Prochlorococcus metapangenome workflow, and we are deeply indebted to their project and their dedication to sharing a reproducible workflows. Our project investiated the environmental representation of the various bacterial pangenomes to learn more about the ecology and evolution of microbial biogeography. -

Discovery of High Affinity Receptors for Dityrosine Through Inverse Virtual Screening and Docking and Molecular Dynamics
Article Discovery of High Affinity Receptors for Dityrosine through Inverse Virtual Screening and Docking and Molecular Dynamics Fangfang Wang 1,*,†, Wei Yang 2,3,† and Xiaojun Hu 1,* 1 School of Life Science, Linyi University, Linyi 276000, China; [email protected] 2 Department of Microbiology, Biomedicine Discovery Institute, Monash University, Clayton, VIC 3800, Australia, [email protected] 3 Arieh Warshel Institute of Computational Biology, the Chinese University of Hong Kong, 2001 Longxiang Road, Longgang District, Shenzhen 518000, China * Corresponding author: [email protected] † These authors contributed equally to this work. Received: 09 December 2018; Accepted: 23 December 2018; Published: date Table S1. Docking affinity scores for cis-dityrosine binding to binding proteins. Target name PDB/UniProtKB Type Affinity (kcal/mol) Galectin-1 1A78/P56217 Lectin -6.2±0.0 Annexin III 1AXN/P12429 Calcium/phospholipid Binding Protein -7.5±0.0 Calmodulin 1CTR/P62158 Calcium Binding Protein -5.8±0.0 Seminal Plasma Protein Pdc-109 1H8P/P02784 Phosphorylcholine Binding Protein -6.6±0.0 Annexin V 1HAK/P08758 Calcium/phospholipid Binding -7.4±0.0 Alpha 1 antitrypsin 1HP7/P01009 Protein Binding -7.6±0.0 Histidine-Binding Protein 1HSL/P0AEU0 Binding Protein -6.3±0.0 Intestinal Fatty Acid Binding Protein 1ICN/P02693 Binding Protein(fatty Acid) -9.1±0.0* Migration Inhibitory Factor-Related Protein 14 1IRJ/P06702 Metal Binding Protein -7.0±0.0 Lysine-, Arginine-, Ornithine-Binding Protein 1LST/P02911 Amino Acid Binding Protein -6.5±0.0 -

Transient Expression for Engineering Triterpenoid Diversity in Plants
Transient expression for engineering triterpenoid diversity in plants James Ashley Reed A thesis submitted to the University of East Anglia for the degree of Doctor of Philosophy © September 2016 John Innes Centre Norwich Research Park Norwich Norfolk United Kingdom © This copy of the thesis has been supplied on condition that anyone who consults it is understood to recognise that its copyright rests with the author and that use of any information derived there from must be in accordance with current UK Copyright Law. In addition, any quotation or extract must include full attribution. Abstract The triterpenes are one of the largest and most structurally diverse classes of plant specialised metabolites, with important commercial, agronomical and medical potential. However the structural complexity and low abundance of these compounds in nature presents significant challenges for exploiting this diversity. In recent years, advances in our understanding of triterpene biosynthesis in various plant species have greatly facilitated the ability to produce these compounds in other hosts. Plant-based host systems hold great promise because they may provide substrates, cofactors and subcellular compartmentalisation mechanisms facilitating engineering production of complex triterpenoids. Agroinfiltration is a rapid and scalable process that enables transient expression of biosynthetic genes in amenable plants such as Nicotiana benthamiana. This technique holds great promise for selective production and modification of triterpenes, but studies in this area have been limited. In this PhD thesis, transient expression is used to produce and oxidise triterpenes in N. benthamiana. Triterpene yields are improved through expression of rate-limiting genes in the mevalonate pathway (Chapter 3). An enzymatic ‘toolkit’ is established from a collection of previously characterised triterpene biosynthetic genes, allowing production of milligram quantities of a diverse array of simple and oxidised triterpenes in N. -

Engineering Metabolic Systems for Production of Advanced Fuels
J Ind Microbiol Biotechnol (2009) 36:471–479 DOI 10.1007/s10295-009-0532-0 REVIEW Engineering metabolic systems for production of advanced fuels Yajun Yan · James C. Liao Received: 7 December 2008 / Accepted: 14 January 2009 / Published online: 7 February 2009 © The Author(s) 2009. This article is published with open access at Springerlink.com Abstract The depleting petroleum storage and increasing limitations, such as low energy density, high vapor environmental deterioration are threatening the sustainable pressure, and corrosiveness, which prevent its widespread development of human societies. As such, biofuels and utilization given the existing infrastructure. chemical feedstocks generated from renewable sources are Higher alcohols (with more than two carbons), biodie- becoming increasingly important. Although previous eVorts sels, and fatty acid derivatives are thought to be more suit- led to great success in bio-ethanol production, higher alco- able fuels. Their physicochemical properties are more hols, fatty acid derivatives including biodiesels, alkanes, compatible with gasoline-based fuels and allow direct utili- and alkenes oVer additional advantages because of their zation of existing infrastructure for storage and distribution. compatibility with existing infrastructure. In addition, some Furthermore, some of these fuel molecules also serve as of these compounds are useful chemical feedstocks. Since important chemical feedstocks. Although the individual native organisms do not naturally produce these com- biochemical steps for synthesizing these compounds in pounds in high quantities, metabolic engineering becomes microbes have been described previously, eVorts in putting essential in constructing producing organisms. In this arti- together highly productive metabolic systems have only cle, we brieXy review the four major metabolic systems, the begun recently.