Implications for Management of the Yampa River Through Dinosaur National Monument Final Report Submitted to the National Park Service
Total Page:16
File Type:pdf, Size:1020Kb

Load more
Recommended publications
-

Operation of Flaming Gorge Dam Final Environmental Impact Statement
Record of Decision Operation of Flaming Gorge Dam Final Environmental Impact Statement I. Summary of Action and Background The Bureau of Reclamation (Reclamation) has completed a final environmental impact statement (EIS) on the operation of Flaming Gorge Dam. The EIS describes the potential effects of modifying the operation of Flaming Gorge Dam to assist in the recovery of four endangered fish, and their critical habitat, downstream from the dam. The four endangered fish species are Colorado pikeminnow (Ptychocheilus lucius), humpback chub (Gila cypha), razorback sucker (Xyrauchen texanus), and bonytail (Gila elegans). Reclamation would implement the proposed action by modifying the operations of Flaming Gorge Dam, to the extent possible, to achieve the flows and temperatures recommended by participants of the Upper Colorado River Endangered Fish Recovery Program (Recovery Program). Reclamation’s goal is to implement the proposed action and, at the same time, maintain and continue all authorized purposes of the Colorado River Storage Project. The purpose of the proposed action is to operate Flaming Gorge Dam to protect and assist in recovery of the populations and designated critical habitat of the four endangered fishes, while maintaining all authorized purposes of the Flaming Gorge Unit of the Colorado River Storage Project (CRSP), including those related to the development of water resources in accordance with the Colorado River Compact. As the Federal agency responsible for the operation of Flaming Gorge Dam, Reclamation was the lead agency in preparing the EIS. Eight cooperating agencies also participated in preparing this EIS: the Bureau of Indian Affairs (BIA), Bureau of Land Management, National Park Service, State of Utah Department of Natural Resources, U.S. -

Big-River Monitoring on the Colorado Plateau
I NVENTORY & M ONITORING N ETWORK Big-River Monitoring on the Colorado Plateau Dustin Perkins1, Mike Scott2, Greg Auble2, Mark Wondzell3, Chris Holmquist-Johnson2, Eric Wahlig2, Helen Thomas1, and Aneth Wight1; 1Northern Colorado Plateau Network, P.O. Box 848, Bldg. 11, Arches National Park, Moab, UT 84532 2U.S. Geological Survey, Biological Resources Discipline, FORT Science Center, 2150 Centre Ave., Building C, Fort Collins, CO 80526; 3National Park Service, Water Resources Division, 1201 Oakridge Dr., Ste. 150, Fort Collins, CO 80525 Introduction and Green rivers in Canyonlands National Park. The Yampa River is the longest relatively free-flowing river Water has always been in short supply in the western reach remaining in the Colorado River basin. The U.S., making it a consistent source of conflict. In Green River is highly regulated by Flaming Gorge Dam the Colorado River drainage, an increasing human but is partially restored below its confluence with the population fuels increased demands for water from Yampa River. There have been large-scale changes the river and its tributaries. As a result, streamflow to the Green River since Flaming Gorge Dam was in virtually all of these systems has been altered by completed in 1962. reservoirs and other water-development projects. In most cases, reduced flows have significantly altered Monitoring of these rivers and their riparian peak flows and increased base flows that structure vegetation focuses on processes that affect the river floodplain vegetation, stream-channel morphology, channel, active bars, and riparian floodplains. To get and water quality (e.g., temperature, suspended a complete picture of river conditions, the NCPN sediment, nutrients). -

River Flow Advisory
River Flow Advisory Bureau . of Reclamation Upper Colorado Region Salt Lake City, Utah Vol. 15, No. 1 September 1984 River flows in the Upper Colorado River drainage, still high for this time of year, are not expected to decrease much for several weeks: While the daily update of operations and releases has been discontinued, the toll-free numbers now provide updates on Bureau of Reclamation activities and projects. Utah residents may call 1-800-624-1094 and out-of-Utah residents may call 1-800-624-5099. Colorado River at Westwater Canyon The flow of the Colorado River on September 10 was 7, 000 cfs, and is expected to decrease slightly over the next few weeks. Cataract Canyon Includin2 the Green- River The flow was 11,500 cfs on September 10 and will continue to decrease slightly. Lake Powell Lake Powell's elevation on September 10 was 3,699. Assuming normal inflow for this time of year, the lake should continue to go down slowly to elevation 3,682 by next spring. Colorado River through Grand Canyon . Releases through Glen Canyon Dam remain at 25,000 cfs. These releases are expected to be maintained with no daily fluctuations in river flows. Upper Green River - Fontenelle Reservoir Fontenelle Reservoir is now at elevation 6,482 feet. Releases through the dam will be reduced to about 600 cfs starting on September 17 for about 2 weeks during powerplant maintenance. Green River Flows Below Flaming Gorge Dam On September 10 Flaming Gorge Reservoir was at elevation 6,039.9 feet. Releases from the dam are expected to average 2, 500 cfs in September and October with usual daily fluctuations. -

Gunnison River
final environmental statement wild and scenic river study september 1979 GUNNISON RIVER COLORADO SPECIAL NOTE This environmental statement was initiated by the Bureau of Outdoor Recreation (BOR) and the Colorado Department of Natural Resources in January, 1976. On January 30, 1978, a reorganization within the U.S. Department of the Interior resulted in BOR being restructured and renamed the Heritage Conservation and Recreation Service (HCRS). On March 27, 1978, study responsibility was transferred from HCRS to the National Park Service. The draft environmental statement was prepared by HCRS and cleared by the U.S. Department of the Interior prior to March 27, 1978. Final revisions and publication of both the draft environmental statement, as well as this document have been the responstbility of the National Park Service. FINAL ENVIRONMENTAL STATEMENT GUNNISON WILD AND SCENIC RIVER STUDY Prepared by United States Department of the Interior I National Park Service in cooperation with the Colorado Department of Natural Resources represented by the Water Conservation Board staff Director National Par!< Service SUMMARY ( ) Draft (X) Final Environmental Statement Department of the Interior, National Park Service 1. Type of action: ( ) Administrative (X) Legislative 2. Brief description of action: The Gunnison Wild and Scenic River Study recommends inclusion of a 26-mile (41.8-km) segment of the Gunnison River, Colorado, and 12,900 acres (S,200 ha) of adjacent land to be classified as wild in the National Wild and Scenic Rivers System under the administration of the National Park Service and the Bureau of Land Management, U.S. D. I. This river segment extends from the upstream boundary of the Black Canyon of the Gunnison National Monument to approximately 1 mile (1.6 km) below the confluence with the Smith Fork. -

Itinerary: the Yampa River: 5 Days/4 Nights
Itinerary: PO Box 1324 Moab, UT 84532 (800) 332-2439 The Yampa River: (435) 259-8229 Fax (435) 259-2226 Email: [email protected] 5 Days/4 Nights www.GriffithExp.com T h r o u g h Dinosaur National Monument O v e r v i e w of The Yampa River Meeting Place Best Western Antlers 423 West Main Street Vernal, UT 84078 Meeting Time : 6 : 3 0 pm (MDT) The evening before your trip Orientation: 6 : 3 0 pm (MDT) the day BEFORE d e p a r t u r e H e r e you will learn what to expect and prepare for, receive your dry bags, sign Assumption of Risk forms, and get a chance to ask last minute q u e s t i o n s . Morning Place : Best Western Antlers 423 West Main Street Vernal, UT 84078 M o r n i n g T i m e : 7 : 0 0 a m (MDT) Return Time : Approximately 5 : 0 0 - 6 : 0 0 P M Rapid Rating: C l a s s I I I - I V (water level dependent) # of Rapids : 16 River Miles: 72 P u t i n : Deer Lodge Park Ranger Station T a k e - out : Split Mountain boat ramp Trip Length: 5 D a y s / 4 N i g h t s Raft Type(s): O a r b o a t s , Paddleboats and Inflatable Kayaks Age Limit: Minimum Age is 10 y e a r s o l d What makes this trip special? The Yampa River through the Dinosaur National Monument has it all! As the last free-flowing river in the entire Colorado River drainage, the Yampa is incredibly wild in May and June. -

Executive Summary U.S
Glen Canyon Dam Long-Term Experimental and Management Plan Environmental Impact Statement PUBLIC DRAFT Executive Summary U.S. Department of the Interior Bureau of Reclamation, Upper Colorado Region National Park Service, Intermountain Region December 2015 Cover photo credits: Title bar: Grand Canyon National Park Grand Canyon: Grand Canyon National Park Glen Canyon Dam: T.R. Reeve High-flow experimental release: T.R. Reeve Fisherman: T. Gunn Humpback chub: Arizona Game and Fish Department Rafters: Grand Canyon National Park Glen Canyon Dam Long-Term Experimental and Management Plan December 2015 Draft Environmental Impact Statement 1 CONTENTS 2 3 4 ACRONYMS AND ABBREVIATIONS .................................................................................. vii 5 6 ES.1 Introduction ............................................................................................................ 1 7 ES.2 Proposed Federal Action ........................................................................................ 2 8 ES.2.1 Purpose of and Need for Action .............................................................. 2 9 ES.2.2 Objectives and Resource Goals of the LTEMP ....................................... 3 10 ES.3 Scope of the DEIS .................................................................................................. 6 11 ES.3.1 Affected Region and Resources .............................................................. 6 12 ES.3.2 Impact Topics Selected for Detailed Analysis ........................................ 6 13 ES.4 -

Management of the Colorado River: Water Allocations, Drought, and the Federal Role
Management of the Colorado River: Water Allocations, Drought, and the Federal Role Updated March 21, 2019 Congressional Research Service https://crsreports.congress.gov R45546 SUMMARY R45546 Management of the Colorado River: Water March 21, 2019 Allocation, Drought, and the Federal Role Charles V. Stern The Colorado River Basin covers more than 246,000 square miles in seven U.S. states Specialist in Natural (Wyoming, Colorado, Utah, New Mexico, Arizona, Nevada, and California) and Resources Policy Mexico. Pursuant to federal law, the Bureau of Reclamation (part of the Department of the Interior) manages much of the basin’s water supplies. Colorado River water is used Pervaze A. Sheikh primarily for agricultural irrigation and municipal and industrial (M&I) uses, but it also Specialist in Natural is important for power production, fish and wildlife, and recreational uses. Resources Policy In recent years, consumptive uses of Colorado River water have exceeded natural flows. This causes an imbalance in the basin’s available supplies and competing demands. A drought in the basin dating to 2000 has raised the prospect of water delivery curtailments and decreased hydropower production, among other things. In the future, observers expect that increasing demand for supplies, coupled with the effects of climate change, will further increase the strain on the basin’s limited water supplies. River Management The Law of the River is the commonly used shorthand for the multiple laws, court decisions, and other documents governing Colorado River operations. The foundational document of the Law of the River is the Colorado River Compact of 1922. Pursuant to the compact, the basin states established a framework to apportion the water supplies between the Upper and Lower Basins of the Colorado River, with the dividing line between the two basins at Lee Ferry, AZ (near the Utah border). -

Navajo Reservoir and San Juan River Temperature Study 2006
NAVAJO RESERVOIR AND SAN JUAN RIVER TEMPERATURE STUDY NAVAJO RESERVOIR BUREAU OF RECLAMATION 125 SOUTH STATE STREET SALT LAKE CITY, UT 84138 Navajo Reservoir and San Juan River Temperature Study Page ii NAVAJO RESERVOIR AND SAN JUAN RIVER TEMPERATURE STUDY PREPARED FOR: SAN JUAN RIVER ENDANGERED FISH RECOVERY PROGRAM BY: Amy Cutler U.S. Department of the Interior Bureau of Reclamation Upper Colorado Regional Office FINAL REPORT SEPTEMBER 1, 2006 ii Navajo Reservoir and San Juan River Temperature Study Page iii TABLE OF CONTENTS EXECUTIVE SUMMARY ...............................................................................................1 1. INTRODUCTION......................................................................................................3 2. OBJECTIVES ............................................................................................................5 3. MODELING OVERVIEW .......................................................................................6 4. RESERVOIR TEMPERATURE MODELING ......................................................7 5. RIVER TEMPERATURE MODELING...............................................................14 6. UNSTEADY RIVER TEMPERATURE MODELING........................................18 7. ADDRESSING RESERVOIR SCENARIOS USING CE-QUAL-W2................23 7.1 Base Case Scenario............................................................................................23 7.2 TCD Scenarios...................................................................................................23 -

Flaming Gorge Operation Plan - May 2021 Through April 2022
Flaming Gorge Operation Plan - May 2021 through April 2022 Concurrence by Kathleen Callister, Resources Management Division Manager Kent Kofford, Provo Area Office Manager Nicholas Williams, Upper Colorado Basin Power Manager Approved by Wayne Pullan, Upper Colorado Basin Regional Director U.S. Department of the Interior Bureau of Reclamation • Interior Region 7 Upper Colorado Basin • Power Office Salt Lake City, Utah May 2021 Purpose This Flaming Gorge Operation Plan (FG-Ops) fulfills the 2006 Flaming Gorge Record of Decision (ROD) requirement for May 2021 through April 2022. The FG-Ops also completes the 4-step process outlined in the Flaming Gorge Standard Operation Procedures. The Upper Colorado Basin Power Office (UCPO) operators will fulfil the operation plan and may alter from FG-Ops due to day to day conditions, although we will attempt to stay within the boundaries of the operations defined below. Listed below are proposed operation plans for four different scenarios: moderately dry, average (above median), average (below median), and moderately wet. As of the publishing of this document, the most likely scenario is the moderately dry, however actual operations will vary with hydrologic conditions. The Upper Colorado River Endangered Fish Recovery Program (Recovery Program), the Flaming Gorge Technical Working Group (FGTWG), Flaming Gorge Working Group (FG WG), United States Fish and Wildlife Service (FWS) and Western Area Power Administration (WAPA) provided input that was considered in the development of this report. The FG-Ops describes the current hydrologic classification of the Green River Basin and the hydrologic conditions in the Yampa River Basin. The FG-Ops identifies the most likely Reach 2 peak flow magnitude and duration that is to be targeted for the upcoming spring flows. -
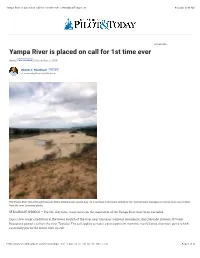
Yampa River Is Placed on Call for 1St Time Ever | Steamboattoday.Com 8/12/20, 9�43 AM
Yampa River is placed on call for 1st time ever | SteamboatToday.com 8/12/20, 943 AM YOUR AD HERE » Yampa River is placed on call for 1st time ever News FOLLOW NEWS | September 5, 2018 Eleanor C. Hasenbeck FOLLOW [email protected] The Yampa River flows through Dinosaur National Monument around Aug. 18. Low flows in the lower stretch of the river led water managers to curtail some use of water from the river. (courtesy photo) STEAMBOAT SPRINGS — For the first time, water users on the main stem of the Yampa River have been curtailed. Due to low water conditions in the lower stretch of the river near Dinosaur National Monument, the Colorado Division of Water Resources placed a call on the river Tuesday. The call applies to water users upstream from the river’s lowest diversion point, which essentially places the entire river on call. https://www.steamboatpilot.com/news/yampa-river-is-placed-on-call-for-1st-time-ever/ Page 1 of 4 Yampa River is placed on call for 1st time ever | SteamboatToday.com 8/12/20, 943 AM “We are now faced again with the Yampa River being extremely low at its lower end, and we are unable to both protect the Endangered Fishes Recovery Program reservoir water and allow all water users to continue to divert water,” Erin Light, the area division engineer, wrote in an email to water users in the Yampa River Basin on Wednesday morning. The Division of Water Resources places a call on a stream when water rights owners do not receive the amount of water they have a legal right to. -

Flaming Gorge Operation Plan - May 2020 Through April 2021
Flaming Gorge Operation Plan - May 2020 through April 2021 Concurrence by Date Kathleen Callister, Resource Management Division Chief Date ___________, Provo Area Office Manager Date ___________, Upper Colorado Power Manager Approved by Date Brent Esplin, Upper Colorado Regional Director Purpose Blue for changes that will happen This Flaming Gorge Operation Plan (FG-Ops) fulfills the 2006 Flaming Gorge Record of Decision (ROD) requirement for May 2020 through April 2021. The FG-Ops also completes the 4-step process outlined in the Flaming Gorge Standard Operation Procedures. The Upper Colorado Basin Power Office (UCPO) operators will fulfil the operation plan and may alter from FG-Ops due to day to day conditions, although we will attempt to stay within the boundaries of the operations defined below. Listed below are proposed operation plans for 3 different scenarios: moderately dry, average (above / below median), and moderately wet. As of the publishing of this document, the most likely scenario is the average (below median), however actual operations will vary with hydrologic conditions. This operation plan covers Flaming Gorge operation from May 2020 through April 2021 per the Flaming Gorge Final Environmental Impact (EIS). Both the Upper Colorado River Endangered Fish Recovery Program (Recovery Program) and the Flaming Gorge Technical Working Group (FGTWG) along with input from the Flaming Gorge Working Group, United States Fish and Wildlife Service (FWS) and Western Area Power Administration (WAPA) provided input that is incorporated into the Final FG-Ops. The 2005 Final EIS specifically addresses and outlines the content of the FGTWG proposal. This outline is used for the basis of the FG-Ops. -

CRWAS Phase I Yampa River – Statemod Model Brief
CRWAS Phase I Yampa River – StateMod Model Brief 1. Introduction................................................................................................................................. 2 2. Model Components..................................................................................................................... 4 2.1 Inflow Hydrology.................................................................................................................4 2.1.1 Data Sources and Filling Techniques.........................................................................6 2.2 Physical Systems.................................................................................................................. 8 2.2.1 Data Sources and Filling Techniques.........................................................................9 2.3 Water Demands...................................................................................................................10 2.3.1 Data Sources and Filling Techniques...................................................................... 10 2.4 Legal and Administrative Conditions................................................................................ 11 2.4.1 Data Sources and Filling Techniques.......................................................................12 3. Model Calibration......................................................................................................................13 3.1 First Step Calibration..........................................................................................................13