The Problem of Vertebrate Ancestry, 1859-1875
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Sahel Journal of Veterinary Sciences
Original Article Sahel Journal of Veterinary Sciences Article History Received: 4th March, 2020 Sahel J. Vet. Sci. Vol. 17, No. 2, pp. 19-25 (2020) Revised: 3rd June, 2020 Copyright © 2020 Faculty of Veterinary Medicine, University of Maiduguri Accepted: 7th June, 2020 All rights reserved Published: 30th June, 2020 Structure of the Leydig Cell in the African Sideneck Turtle (Pelusios castaneus) Olukole, S. G. and Oke, B. O. Department of Veterinary Anatomy, Faculty of Veterinary Medicine, University of Ibadan, Ibadan, Nigeria. * Author for Correspondence: [email protected]; +2348033574752 Abstract The African sideneck turtle (Pelusios castaneus) is a freshwater turtle of West African origin used in traditional medicine with little consumption as meat. There have been documentations on the reproductive biology of the turtle with no report on the structure of the Leydig cell of the animal. We described the structure of the Leydig cell of the adult African sideneck turtle using histology, microstereology and transmission electron microscopy. The Leydig cell of the African sideneck turtle were elliptical in shape when found proximal to blood vessels and elongated at other points within the testicular interstitium. Leydig cells occurred in cords or clusters of varying sizes and numbers (3-5 cells) that appear to be random in distribution possessing round to ovoid nuclei containing small amount of peripherally disposed heterochromatin with prominent nucleoli. The seminiferous tubules of the turtle occupied about 85% of the total testicular parenchyma while the interstitium occupied 15% of it. Of this 15%, the Leydig cell occupied about 10% while the stromal elements, inclusive of blood vessels occupied the remaining 5%. -

I FLATWORM PREDATION on JUVENILE FRESHWATER
FLATWORM PREDATION ON JUVENILE FRESHWATER MUSSELS A Thesis Presented to the Graduate College of Southwest Missouri State University In Partial Fulfillment of the Requirements for the Master of Science Degree By Angela Marie Delp July 2002 i FLATWORM PREDATION OF JUVENILE FRESHWATER MUSSELS Biology Department Southwest Missouri State University, July 27, 2002 Master of Science in Biology Angela Marie Delp ABSTRACT Free-living flatworms (Phylum Platyhelminthes, Class Turbellaria) are important predators on small aquatic invertebrates. Macrostomum tuba, a predominantly benthic species, feeds on juvenile freshwater mussels in fish hatcheries and mussel culture facilities. Laboratory experiments were performed to assess the predation rate of M. tuba on newly transformed juveniles of plain pocketbook mussel, Lampsilis cardium. Predation rate at 20 oC in dishes without substrate was 0.26 mussels·worm-1·h-1. Predation rate increased to 0.43 mussels·worm-1·h-1 when a substrate, polyurethane foam, was present. Substrate may have altered behavior of the predator and brought the flatworms in contact with the mussels more often. An alternative prey, the cladoceran Ceriodaphnia reticulata, was eaten at a higher rate than mussels when only one prey type was present, but at a similar rate when both were present. Finally, the effect of flatworm size (0.7- 2.2 mm long) on predation rate on mussels (0.2 mm) was tested. Predation rate increased with predator size. The slope of this relationship decreased with increasing predator size. Predation rate was near zero in 0.7 mm worms. Juvenile mussels grow rapidly and can escape flatworm predation by exceeding the size of these tiny predators. -

The Physiology and Immunology of the Endocrine Testis
tì/ jì;:llilul s\ t: 1.q_ 81 LI Àt{Y THE PHYSIOLOGY AND IMMUNOLOGY OF THE ENDOCRINE TESTIS by Simon MADDOCKS, B.Ag.Scl. (Hons) A thesls submltted to the Unlversfty of Adelalde ln fulfl'lment of the requlrements for the degree of Doctor of PhllosoPhY Department of Anlmal Scfences Walte Agrlcultural Research Instltute The UnlversitY of Adelalde March 1987 lì,.,ì,r(,:,td r/ riÈ"1 Know'ledge and wlsdom far f rom belng oner. Have ofl tlmes no connexion. Knorledge dwells In heads replete with thoughts of other men; Wlsdom ln minds attentlve to thelr otvn' Know'ledger a rude unprof ltab'le massr The meré materials with whlch wlsdom bul'ldsr Tfl.|smoothldandsquarldandflttedtoftsplacer Does but encumber whom lt seems trenrlch' Knowledge ls proud that he has learnrd so much; Wisdom ls humb'le that he knows no more' I{il'l lam CowPer ( 1731-1800) '_ .å Cowper Poetical Works TABLE OF CONTENTS f CONTENTS vii ABSTRACT x DECLARATION xl ACKNOt{LEDGEMENTS PREFACE xlff PART I: T CTIAPTER I: INTRODUCTION 2 1. 1. INTRODUCTION 2 L.2, THE PHYSIOLOGY OF THE ENDOCRINE TESTIS L.2,L. Introducti on 2 L.2.2. The Leydfg cells 7 r.2,2. I, Anatomy 7 L.2.2. 2. Relatiónship to blood vessels, lymph vessels and seminiferous tubul es L2 .2.2 3, Re]atfonshlp to other ce]ls in fnterstlt'lal tissue I5 I 18 ,212 4. Hormones produced by the Leydfg ce1ìs I 22 L .2.3 The Sertoll cells 24 T .2,3 I. AnatomY I .2.3 2. -

Cumulated Bibliography of Biographies of Ocean Scientists Deborah Day, Scripps Institution of Oceanography Archives Revised December 3, 2001
Cumulated Bibliography of Biographies of Ocean Scientists Deborah Day, Scripps Institution of Oceanography Archives Revised December 3, 2001. Preface This bibliography attempts to list all substantial autobiographies, biographies, festschrifts and obituaries of prominent oceanographers, marine biologists, fisheries scientists, and other scientists who worked in the marine environment published in journals and books after 1922, the publication date of Herdman’s Founders of Oceanography. The bibliography does not include newspaper obituaries, government documents, or citations to brief entries in general biographical sources. Items are listed alphabetically by author, and then chronologically by date of publication under a legend that includes the full name of the individual, his/her date of birth in European style(day, month in roman numeral, year), followed by his/her place of birth, then his date of death and place of death. Entries are in author-editor style following the Chicago Manual of Style (Chicago and London: University of Chicago Press, 14th ed., 1993). Citations are annotated to list the language if it is not obvious from the text. Annotations will also indicate if the citation includes a list of the scientist’s papers, if there is a relationship between the author of the citation and the scientist, or if the citation is written for a particular audience. This bibliography of biographies of scientists of the sea is based on Jacqueline Carpine-Lancre’s bibliography of biographies first published annually beginning with issue 4 of the History of Oceanography Newsletter (September 1992). It was supplemented by a bibliography maintained by Eric L. Mills and citations in the biographical files of the Archives of the Scripps Institution of Oceanography, UCSD. -

Number of Living Species in Australia and the World
Numbers of Living Species in Australia and the World 2nd edition Arthur D. Chapman Australian Biodiversity Information Services australia’s nature Toowoomba, Australia there is more still to be discovered… Report for the Australian Biological Resources Study Canberra, Australia September 2009 CONTENTS Foreword 1 Insecta (insects) 23 Plants 43 Viruses 59 Arachnida Magnoliophyta (flowering plants) 43 Protoctista (mainly Introduction 2 (spiders, scorpions, etc) 26 Gymnosperms (Coniferophyta, Protozoa—others included Executive Summary 6 Pycnogonida (sea spiders) 28 Cycadophyta, Gnetophyta under fungi, algae, Myriapoda and Ginkgophyta) 45 Chromista, etc) 60 Detailed discussion by Group 12 (millipedes, centipedes) 29 Ferns and Allies 46 Chordates 13 Acknowledgements 63 Crustacea (crabs, lobsters, etc) 31 Bryophyta Mammalia (mammals) 13 Onychophora (velvet worms) 32 (mosses, liverworts, hornworts) 47 References 66 Aves (birds) 14 Hexapoda (proturans, springtails) 33 Plant Algae (including green Reptilia (reptiles) 15 Mollusca (molluscs, shellfish) 34 algae, red algae, glaucophytes) 49 Amphibia (frogs, etc) 16 Annelida (segmented worms) 35 Fungi 51 Pisces (fishes including Nematoda Fungi (excluding taxa Chondrichthyes and (nematodes, roundworms) 36 treated under Chromista Osteichthyes) 17 and Protoctista) 51 Acanthocephala Agnatha (hagfish, (thorny-headed worms) 37 Lichen-forming fungi 53 lampreys, slime eels) 18 Platyhelminthes (flat worms) 38 Others 54 Cephalochordata (lancelets) 19 Cnidaria (jellyfish, Prokaryota (Bacteria Tunicata or Urochordata sea anenomes, corals) 39 [Monera] of previous report) 54 (sea squirts, doliolids, salps) 20 Porifera (sponges) 40 Cyanophyta (Cyanobacteria) 55 Invertebrates 21 Other Invertebrates 41 Chromista (including some Hemichordata (hemichordates) 21 species previously included Echinodermata (starfish, under either algae or fungi) 56 sea cucumbers, etc) 22 FOREWORD In Australia and around the world, biodiversity is under huge Harnessing core science and knowledge bases, like and growing pressure. -

PALEONTOLOGICAL TECHNICAL REPORT: 6Th AVENUE and WADSWORTH BOULEVARD INTERCHANGE PHASE II ENVIRONMENTAL ASSESSMENT, CITY of LAKEWOOD, JEFFERSON COUNTY, COLORADO
PALEONTOLOGICAL TECHNICAL REPORT: 6th AVENUE AND WADSWORTH BOULEVARD INTERCHANGE PHASE II ENVIRONMENTAL ASSESSMENT, CITY OF LAKEWOOD, JEFFERSON COUNTY, COLORADO Prepared for: TEC Inc. 1746 Cole Boulevard, Suite 265 Golden, CO 80401 Prepared by: Paul C. Murphey, Ph.D. and David Daitch M.S. Rocky Mountain Paleontology 4614 Lonespur Court Oceanside, CA 92056 303-514-1095; 760-758-4019 www.rockymountainpaleontology.com Prepared under State of Colorado Paleontological Permit 2007-33 January, 2007 TABLE OF CONTENTS 1.0 SUMMARY............................................................................................................................. 3 2.0 INTRODUCTION ................................................................................................................... 4 2.1 DEFINITION AND SIGNIFICANCE OF PALEONTOLOGICAL RESOURCES........... 4 3.0 METHODS .............................................................................................................................. 6 4.0. LAWS, ORDINANCES, REGULATIONS AND STANDARDS......................................... 7 4.1. Federal................................................................................................................................. 7 4.2. State..................................................................................................................................... 8 4.3. County................................................................................................................................. 8 4.4. City..................................................................................................................................... -
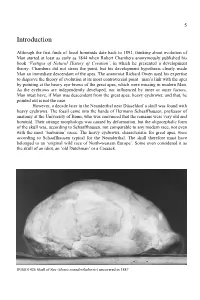
Introduction
5 Introduction Although the first finds of fossil hominids date back to 1891, thinking about evolution of Man started at least as early as 1844 when Robert Chambers anonymously published his book ‘Vestiges of Natural History of Creation’, in which he presented a development theory. Chambers did not stress the point, but his development hypothesis clearly made Man an immediate descendant of the apes. The anatomist Richard Owen used his expertise to disprove the theory of evolution at its most controversial point –man’s link with the apes by pointing at the heavy eye-brows of the great apes, which were missing in modern Man. As the eyebrows are independently developed, nor influenced by inner or outer factors, Man must have, if Man was descendent from the great apes, heavy eyebrows; and that, he pointed out is not the case. However, a decade later in the Neanderthal near Düsseldorf a skull was found with heavy eyebrows. The fossil came into the hands of Hermann Schaaffhausen, professor of anatomy at the University of Bonn, who was convinced that the remains were very old and hominid. Their strange morphology was caused by deformation, but the oligocephalic form of the skull was, according to Schaaffhausen, not comparable to any modern race, not even with the most ‘barbarian’ races. The heavy eyebrows, characteristic for great apes, were according to Schaaffhausen typical for the Neanderthal. The skull therefore must have belonged to an ‘original wild race of North-western Europe’. Some even considered it as the skull of an idiot, an ‘old Dutchman’ or a Cossack. -

Amphibian Alliance for Zero Extinction Sites in Chiapas and Oaxaca
Amphibian Alliance for Zero Extinction Sites in Chiapas and Oaxaca John F. Lamoreux, Meghan W. McKnight, and Rodolfo Cabrera Hernandez Occasional Paper of the IUCN Species Survival Commission No. 53 Amphibian Alliance for Zero Extinction Sites in Chiapas and Oaxaca John F. Lamoreux, Meghan W. McKnight, and Rodolfo Cabrera Hernandez Occasional Paper of the IUCN Species Survival Commission No. 53 The designation of geographical entities in this book, and the presentation of the material, do not imply the expression of any opinion whatsoever on the part of IUCN concerning the legal status of any country, territory, or area, or of its authorities, or concerning the delimitation of its frontiers or boundaries. The views expressed in this publication do not necessarily reflect those of IUCN or other participating organizations. Published by: IUCN, Gland, Switzerland Copyright: © 2015 International Union for Conservation of Nature and Natural Resources Reproduction of this publication for educational or other non-commercial purposes is authorized without prior written permission from the copyright holder provided the source is fully acknowledged. Reproduction of this publication for resale or other commercial purposes is prohibited without prior written permission of the copyright holder. Citation: Lamoreux, J. F., McKnight, M. W., and R. Cabrera Hernandez (2015). Amphibian Alliance for Zero Extinction Sites in Chiapas and Oaxaca. Gland, Switzerland: IUCN. xxiv + 320pp. ISBN: 978-2-8317-1717-3 DOI: 10.2305/IUCN.CH.2015.SSC-OP.53.en Cover photographs: Totontepec landscape; new Plectrohyla species, Ixalotriton niger, Concepción Pápalo, Thorius minutissimus, Craugastor pozo (panels, left to right) Back cover photograph: Collecting in Chamula, Chiapas Photo credits: The cover photographs were taken by the authors under grant agreements with the two main project funders: NGS and CEPF. -

Phylum Arthropod Silvia Rondon, and Mary Corp, OSU Extension Entomologist and Agronomist, Respectively Hermiston Research and Extension Center, Hermiston, Oregon
Phylum Arthropod Silvia Rondon, and Mary Corp, OSU Extension Entomologist and Agronomist, respectively Hermiston Research and Extension Center, Hermiston, Oregon Member of the Phyllum Arthropoda can be found in the seas, in fresh water, on land, or even flying freely; a group with amazing differences of structure, and so abundant that all the other animals taken together are less than 1/6 as many as the arthropods. Well-known members of this group are the Kingdom lobsters, crayfish and crabs; scorpions, spiders, mites, ticks, Phylum Phylum Phylum Class the centipedes and millipedes; and last, but not least, the Order most abundant of all, the insects. Family Genus The Phylum Arthropods consist of the following Species classes: arachnids, chilopods, diplopods, crustaceans and hexapods (insects). All arthropods possess: • Exoskeleton. A hard protective covering around the outside of the body (divided by sutures into plates called sclerites). An insect's exoskeleton (integument) serves as a protective covering over the body, but also as a surface for muscle attachment, a water-tight barrier against desiccation, and a sensory interface with the environment. It is a multi-layered structure with four functional regions: epicuticle (top layer), procuticle, epidermis, and basement membrane. • Segmented body • Jointed limbs and jointed mouthparts that allow extensive specialization • Bilateral symmetry, whereby a central line can divide the body Insect molting or removing its into two identical halves, left and right exoesqueleton • Ventral nerve -

The Correspondence of Julius Haast and Joseph Dalton Hooker, 1861-1886
The Correspondence of Julius Haast and Joseph Dalton Hooker, 1861-1886 Sascha Nolden, Simon Nathan & Esme Mildenhall Geoscience Society of New Zealand miscellaneous publication 133H November 2013 Published by the Geoscience Society of New Zealand Inc, 2013 Information on the Society and its publications is given at www.gsnz.org.nz © Copyright Simon Nathan & Sascha Nolden, 2013 Geoscience Society of New Zealand miscellaneous publication 133H ISBN 978-1-877480-29-4 ISSN 2230-4495 (Online) ISSN 2230-4487 (Print) We gratefully acknowledge financial assistance from the Brian Mason Scientific and Technical Trust which has provided financial support for this project. This document is available as a PDF file that can be downloaded from the Geoscience Society website at: http://www.gsnz.org.nz/information/misc-series-i-49.html Bibliographic Reference Nolden, S.; Nathan, S.; Mildenhall, E. 2013: The Correspondence of Julius Haast and Joseph Dalton Hooker, 1861-1886. Geoscience Society of New Zealand miscellaneous publication 133H. 219 pages. The Correspondence of Julius Haast and Joseph Dalton Hooker, 1861-1886 CONTENTS Introduction 3 The Sumner Cave controversy Sources of the Haast-Hooker correspondence Transcription and presentation of the letters Acknowledgements References Calendar of Letters 8 Transcriptions of the Haast-Hooker letters 12 Appendix 1: Undated letter (fragment), ca 1867 208 Appendix 2: Obituary for Sir Julius von Haast 209 Appendix 3: Biographical register of names mentioned in the correspondence 213 Figures Figure 1: Photographs -

A Taxonomic Re-Evaluation of the Allium Sanbornii Complex
University of the Pacific Scholarly Commons University of the Pacific Theses and Dissertations Graduate School 1986 A taxonomic re-evaluation of the Allium sanbornii complex Stella Sue Denison University of the Pacific Follow this and additional works at: https://scholarlycommons.pacific.edu/uop_etds Part of the Biology Commons Recommended Citation Denison, Stella Sue. (1986). A taxonomic re-evaluation of the Allium sanbornii complex. University of the Pacific, Thesis. https://scholarlycommons.pacific.edu/uop_etds/2124 This Thesis is brought to you for free and open access by the Graduate School at Scholarly Commons. It has been accepted for inclusion in University of the Pacific Theses and Dissertations by an authorized administrator of Scholarly Commons. For more information, please contact [email protected]. A TAXONOMIC RE-EVALUATION OF THE ALLIUM SANBORNII COMPLEX A Thesis Presented to the Faculty of the Graduate School University of the Pacific In Partial Fulfillment of the Requirements for the Degree Master of Science by Stella S. Denison August 1986 ACKNOWLEDGMENTS Many contributions have been made for my successful completion of this work. Appreciation is extended to: Drs. Dale McNeal, Alice Hunter, and Anne Funkhouser for their advice and assistance during the research and in the preparation of this manuscript, the entire Biology faculty for their, friendship and suggestions, Ginger Tibbens for the typing of this manuscript, and to my husband, Craig, and my children, Amy, Eric and Deborah for their continued support and encouragement. Grateful acknowledgement is made to the curators of the herbaria from which material was borrowed during this investigation. These herbaria are indicated below by the standard abbreviations of Holmgren and Keuken (1974}. -

Complicated Meckel's Diverticulum and Therapeutic Management
Ulusal Cer Derg 2013; 29: 63-66 Original Investigation DOI: 10.5152/UCD.2013.36 Complicated Meckel's diverticulum and therapeutic management Varlık Erol, Tayfun Yoldaş, Samet Cin, Cemil Çalışkan, Erhan Akgün, Mustafa Korkut Objective: This study aimed to investigate the treatment options and compare patient management with the litera- ABSTRACT ture for patients operated on for an acute abdomen who had complications due to inflammation of the Meckel’s diverticulum at our clinics. Material and Methods: This study retrospectively evaluated 14 patients who had been operated on for acute abdo- men and had been diagnosed with Meckel’s diverticulitis (MD) in Ege University Medical Faculty Department of General Surgery, between October 2007 and October 2012. Results: Fourteen patients with a diagnosis of Meckel’s diverticulitis (MD) were retrospectively analyzed. Radiologi- cally, the abdominal computer tomography showed pathologies compatible with mechanical intestinal obstruction, Meckel’s diverticulitis and peridiverticular abscess, as well as detection of free air within the abdomen on direct abdominal X-ray. Among patients diagnosed with complicated Meckel’s diverticuli (obstruction, diverticulitis, perfo- ration) 10 patients had partial small bowel resection and end-to-end anastomosis (71.5%), three patients underwent diverticulum excision (21.4%), and one patient underwent right hemicolectomy+ileotransversostomy (7.1%). Conclusion: Meckel’s diverticulum is a vestigial remnant of an omphalomesenteric channel in the small bowel. It is a real congenital diverticular abnormality that contains all three layers of the small bowel. Surgical excision should be performed if Meckel’s diverticulum is detected in order to avoid incidental complications such as ulcer- ation, bleeding, bowel obstruction, diverticulitis or perforation.