And Power-Velocity Relationships in a Multi-Joint Movement
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Velocity-Corrected Area Calculation SCIEX PA 800 Plus Empower
Velocity-corrected area calculation: SCIEX PA 800 Plus Empower Driver version 1.3 vs. 32 Karat™ Software Firdous Farooqui1, Peter Holper1, Steve Questa1, John D. Walsh2, Handy Yowanto1 1SCIEX, Brea, CA 2Waters Corporation, Milford, MA Since the introduction of commercial capillary electrophoresis (CE) systems over 30 years ago, it has been important to not always use conventional “chromatography thinking” when using CE. This is especially true when processing data, as there are some key differences between electrophoretic and chromatographic data. For instance, in most capillary electrophoresis separations, peak area is not only a function of sample load, but also of an analyte’s velocity past the detection window. In this case, early migrating peaks move past the detection window faster than later migrating peaks. This creates a peak area bias, as any relative difference in an analyte’s migration velocity will lead to an error in peak area determination and relative peak area percent. To help minimize Figure 1: The PA 800 Plus Pharmaceutical Analysis System. this bias, peak areas are normalized by migration velocity. The resulting parameter is commonly referred to as corrected peak The capillary temperature was maintained at 25°C in all area or velocity corrected area. separations. The voltage was applied using reverse polarity. This technical note provides a comparison of velocity corrected The following methods were used with the SCIEX PA 800 Plus area calculations using 32 Karat™ and Empower software. For Empower™ Driver v1.3: both, standard processing methods without manual integration were used to process each result. For 32 Karat™ software, IgG_HR_Conditioning: conditions the capillary Caesar integration1 was turned off. -

The Origins of Velocity Functions
The Origins of Velocity Functions Thomas M. Humphrey ike any practical, policy-oriented discipline, monetary economics em- ploys useful concepts long after their prototypes and originators are L forgotten. A case in point is the notion of a velocity function relating money’s rate of turnover to its independent determining variables. Most economists recognize Milton Friedman’s influential 1956 version of the function. Written v = Y/M = v(rb, re,1/PdP/dt, w, Y/P, u), it expresses in- come velocity as a function of bond interest rates, equity yields, expected inflation, wealth, real income, and a catch-all taste-and-technology variable that captures the impact of a myriad of influences on velocity, including degree of monetization, spread of banking, proliferation of money substitutes, devel- opment of cash management practices, confidence in the future stability of the economy and the like. Many also are aware of Irving Fisher’s 1911 transactions velocity func- tion, although few realize that it incorporates most of the same variables as Friedman’s.1 On velocity’s interest rate determinant, Fisher writes: “Each per- son regulates his turnover” to avoid “waste of interest” (1963, p. 152). When rates rise, cashholders “will avoid carrying too much” money thus prompting a rise in velocity. On expected inflation, he says: “When...depreciation is anticipated, there is a tendency among owners of money to spend it speedily . the result being to raise prices by increasing the velocity of circulation” (p. 263). And on real income: “The rich have a higher rate of turnover than the poor. They spend money faster, not only absolutely but relatively to the money they keep on hand. -

Influence of Angular Velocity of Pedaling on the Accuracy of The
Research article 2018-04-10 - Rev08 Influence of Angular Velocity of Pedaling on the Accuracy of the Measurement of Cyclist Power Abstract Almost all cycling power meters currently available on the The miscalculation may be—even significantly—greater than we market are positioned on rotating parts of the bicycle (pedals, found in our study, for the following reasons: crank arms, spider, bottom bracket/hub) and, regardless of • the test was limited to only 5 cyclists: there is no technical and construction differences, all calculate power on doubt other cyclists may have styles of pedaling with the basis of two physical quantities: torque and angular velocity greater variations of angular velocity; (or rotational speed – cadence). Both these measures vary only 2 indoor trainer models were considered: other during the 360 degrees of each revolution. • models may produce greater errors; The torque / force value is usually measured many times during slopes greater than 5% (the only value tested) may each rotation, while the angular velocity variation is commonly • lead to less uniform rotations and consequently neglected, considering only its average value for each greater errors. revolution (cadence). It should be noted that the error observed in this analysis This, however, introduces an unpredictable error into the power occurs because to measure power the power meter considers calculation. To use the average value of angular velocity means the average angular velocity of each rotation. In power meters to consider each pedal revolution as perfectly smooth and that use this type of calculation, this error must therefore be uniform: but this type of pedal revolution does not exist in added to the accuracy stated by the manufacturer. -

VELOCITY from Our Establishment in 1957, We Have Become One of the Oldest Exclusive Manufacturers of Commercial Flooring in the United States
VELOCITY From our establishment in 1957, we have become one of the oldest exclusive manufacturers of commercial flooring in the United States. As one of the largest privately held mills, our FAMILY-OWNERSHIP provides a heritage of proven performance and expansive industry knowledge. Most importantly, our focus has always been on people... ensuring them that our products deliver the highest levels of BEAUTY, PERFORMANCE and DEPENDABILITY. (cover) Velocity Move, quarter turn. (right) Velocity Move with Pop Rojo and Azul, quarter turn. VELOCITY 3 velocity 1814 style 1814 style 1814 style 1814 color 1603 color 1604 color 1605 position direction magnitude style 1814 style 1814 style 1814 color 1607 color 1608 color 1609 reaction move constant style 1814 color 1610 vector Velocity Vector, quarter turn. VELOCITY 5 where to use kinetex Healthcare Fitness Centers kinetex overview Acute care hospitals, medical Health Clubs/Gyms office buildings, urgent care • Cardio Centers clinics, outpatient surgery • Stationary Weight Centers centers, outpatient physical • Dry Locker Room Areas therapy/rehab centers, • Snack Bars outpatient imaging centers, etc. • Offices Kinetex® is an advanced textile composite flooring that combines key attributes of • Cafeteria, dining areas soft-surface floor covering with the long-wearing performance characteristics of • Chapel Retail / Mercantile hard-surface flooring. Created as a unique floor covering alternative to hard-surface Wholesale / Retail merchants • Computer room products, J+J Flooring’s Kinetex encompasses an unprecedented range of • Corridors • Checkout / cash wrap performance attributes for retail, healthcare, education and institutional environments. • Diagnostic imaging suites • Dressing rooms In addition to its human-centered qualities and highly functional design, Kinetex • Dry physical therapy • Sales floor offers a reduced environmental footprint compared to traditional hard-surface options. -

Basement Flood Mitigation
1 Mitigation refers to measures taken now to reduce losses in the future. How can homeowners and renters protect themselves and their property from a devastating loss? 2 There are a range of possible causes for basement flooding and some potential remedies. Many of these low-cost options can be factored into a family’s budget and accomplished over the several months that precede storm season. 3 There are four ways water gets into your basement: Through the drainage system, known as the sump. Backing up through the sewer lines under the house. Seeping through cracks in the walls and floor. Through windows and doors, called overland flooding. 4 Gutters can play a huge role in keeping basements dry and foundations stable. Water damage caused by clogged gutters can be severe. Install gutters and downspouts. Repair them as the need arises. Keep them free of debris. 5 Channel and disperse water away from the home by lengthening the run of downspouts with rigid or flexible extensions. Prevent interior intrusion through windows and replace weather stripping as needed. 6 Many varieties of sturdy window well covers are available, simple to install and hinged for easy access. Wells should be constructed with gravel bottoms to promote drainage. Remove organic growth to permit sunlight and ventilation. 7 Berms and barriers can help water slope away from the home. The berm’s slope should be about 1 inch per foot and extend for at least 10 feet. It is important to note permits are required any time a homeowner alters the elevation of the property. -

Simple Harmonic Motion
[SHIVOK SP211] October 30, 2015 CH 15 Simple Harmonic Motion I. Oscillatory motion A. Motion which is periodic in time, that is, motion that repeats itself in time. B. Examples: 1. Power line oscillates when the wind blows past it 2. Earthquake oscillations move buildings C. Sometimes the oscillations are so severe, that the system exhibiting oscillations break apart. 1. Tacoma Narrows Bridge Collapse "Gallopin' Gertie" a) http://www.youtube.com/watch?v=j‐zczJXSxnw II. Simple Harmonic Motion A. http://www.youtube.com/watch?v=__2YND93ofE Watch the video in your spare time. This professor is my teaching Idol. B. In the figure below snapshots of a simple oscillatory system is shown. A particle repeatedly moves back and forth about the point x=0. Page 1 [SHIVOK SP211] October 30, 2015 C. The time taken for one complete oscillation is the period, T. In the time of one T, the system travels from x=+x , to –x , and then back to m m its original position x . m D. The velocity vector arrows are scaled to indicate the magnitude of the speed of the system at different times. At x=±x , the velocity is m zero. E. Frequency of oscillation is the number of oscillations that are completed in each second. 1. The symbol for frequency is f, and the SI unit is the hertz (abbreviated as Hz). 2. It follows that F. Any motion that repeats itself is periodic or harmonic. G. If the motion is a sinusoidal function of time, it is called simple harmonic motion (SHM). -

Hub City Powertorque® Shaft Mount Reducers
Hub City PowerTorque® Shaft Mount Reducers PowerTorque® Features and Description .................................................. G-2 PowerTorque Nomenclature ............................................................................................ G-4 Selection Instructions ................................................................................ G-5 Selection By Horsepower .......................................................................... G-7 Mechanical Ratings .................................................................................... G-12 ® Shaft Mount Reducers Dimensions ................................................................................................ G-14 Accessories ................................................................................................ G-15 Screw Conveyor Accessories ..................................................................... G-22 G For Additional Models of Shaft Mount Reducers See Hub City Engineering Manual Sections F & J DOWNLOAD AVAILABLE CAD MODELS AT: WWW.HUBCITYINC.COM Certified prints are available upon request EMAIL: [email protected] • www.hubcityinc.com G-1 Hub City PowerTorque® Shaft Mount Reducers Ten models available from 1/4 HP through 200 HP capacity Manufacturing Quality Manufactured to the highest quality 98.5% standards in the industry, assembled Efficiency using precision manufactured components made from top quality per Gear Stage! materials Designed for the toughest applications in the industry Housings High strength ductile -

Chapter 3 Motion in Two and Three Dimensions
Chapter 3 Motion in Two and Three Dimensions 3.1 The Important Stuff 3.1.1 Position In three dimensions, the location of a particle is specified by its location vector, r: r = xi + yj + zk (3.1) If during a time interval ∆t the position vector of the particle changes from r1 to r2, the displacement ∆r for that time interval is ∆r = r1 − r2 (3.2) = (x2 − x1)i +(y2 − y1)j +(z2 − z1)k (3.3) 3.1.2 Velocity If a particle moves through a displacement ∆r in a time interval ∆t then its average velocity for that interval is ∆r ∆x ∆y ∆z v = = i + j + k (3.4) ∆t ∆t ∆t ∆t As before, a more interesting quantity is the instantaneous velocity v, which is the limit of the average velocity when we shrink the time interval ∆t to zero. It is the time derivative of the position vector r: dr v = (3.5) dt d = (xi + yj + zk) (3.6) dt dx dy dz = i + j + k (3.7) dt dt dt can be written: v = vxi + vyj + vzk (3.8) 51 52 CHAPTER 3. MOTION IN TWO AND THREE DIMENSIONS where dx dy dz v = v = v = (3.9) x dt y dt z dt The instantaneous velocity v of a particle is always tangent to the path of the particle. 3.1.3 Acceleration If a particle’s velocity changes by ∆v in a time period ∆t, the average acceleration a for that period is ∆v ∆v ∆v ∆v a = = x i + y j + z k (3.10) ∆t ∆t ∆t ∆t but a much more interesting quantity is the result of shrinking the period ∆t to zero, which gives us the instantaneous acceleration, a. -
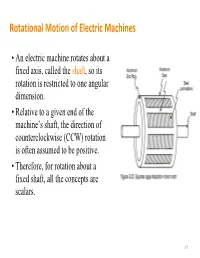
Rotational Motion of Electric Machines
Rotational Motion of Electric Machines • An electric machine rotates about a fixed axis, called the shaft, so its rotation is restricted to one angular dimension. • Relative to a given end of the machine’s shaft, the direction of counterclockwise (CCW) rotation is often assumed to be positive. • Therefore, for rotation about a fixed shaft, all the concepts are scalars. 17 Angular Position, Velocity and Acceleration • Angular position – The angle at which an object is oriented, measured from some arbitrary reference point – Unit: rad or deg – Analogy of the linear concept • Angular acceleration =d/dt of distance along a line. – The rate of change in angular • Angular velocity =d/dt velocity with respect to time – The rate of change in angular – Unit: rad/s2 position with respect to time • and >0 if the rotation is CCW – Unit: rad/s or r/min (revolutions • >0 if the absolute angular per minute or rpm for short) velocity is increasing in the CCW – Analogy of the concept of direction or decreasing in the velocity on a straight line. CW direction 18 Moment of Inertia (or Inertia) • Inertia depends on the mass and shape of the object (unit: kgm2) • A complex shape can be broken up into 2 or more of simple shapes Definition Two useful formulas mL2 m J J() RRRR22 12 3 1212 m 22 JRR()12 2 19 Torque and Change in Speed • Torque is equal to the product of the force and the perpendicular distance between the axis of rotation and the point of application of the force. T=Fr (Nm) T=0 T T=Fr • Newton’s Law of Rotation: Describes the relationship between the total torque applied to an object and its resulting angular acceleration. -

PHYS 211 Lecture 5 - Oscillations: Simple Harmonic Motion 5 - 1
PHYS 211 Lecture 5 - Oscillations: simple harmonic motion 5 - 1 Lecture 5 - Oscillations: simple harmonic motion Text: Fowles and Cassiday, Chap. 3 Consider a power series expansion for the potential energy of an object 2 V(x) = Vo + V1x + V2x + ..., where x is the displacement of an object from its equilibrium position, and Vi, i = 0,1,2... are fixed coefficients. The leading order term in this series is unimportant to the dynamics of the object, since F = -dV/dx and the derivative of a constant vanishes. Further, if we require the equilibrium position x = 0 to be a true minimum in the energy, then V1 = 0. This doesn’t mean that potentials with odd powers in x must vanish, but just says that their minimum is not at x = 0. Thus, the simplest function that one can write is quadratic: 2 V = V2x [A slightly more complicated variant is |x| = (x2)1/2, which is not smooth at x = 0]. For small excursions from equilibrium, the quadratic term is often the leading-order piece of more complex functions such as the Morse or Lennard-Jones potentials. Quadratic potentials correspond to the familiar Hooke’s Law of ideal springs V(x) = kx 2/2 => F(x) = -dV/dx = -(2/2)kx = -kx, where k is the spring constant. Objects subject to Hooke’s Law exhibit oscillatory motion, as can be seen by solving the differential equation arising from Newton’s 2nd law: F = ma => -kx = m(d 2x/dt 2) or d 2x /dt 2 + (k/m)x = 0. We solved this equation in PHYS 120: x(t) = A sin( ot + o) Other functional forms such as cosine can be changed into this form using a suitable choice of the phase angle o. -

Determining the Load Inertia Contribution from Different Power Consumer Groups
energies Article Determining the Load Inertia Contribution from Different Power Consumer Groups Henning Thiesen * and Clemens Jauch Wind Energy Technology Institute (WETI), Flensburg University of Applied Sciences, 24943 Flensburg, Germany; clemens.jauch@hs-flensburg.de * Correspondence: henning.thiesen@hs-flensburg.de Received: 27 February 2020; Accepted: 25 March 2020 ; Published: 1 April 2020 Abstract: Power system inertia is a vital part of power system stability. The inertia response within the first seconds after a power imbalance reduces the velocity of which the grid frequency changes. At present, large shares of power system inertia are provided by synchronously rotating masses of conventional power plants. A minor part of power system inertia is supplied by power consumers. The energy system transformation results in an overall decreasing amount of power system inertia. Hence, inertia has to be provided synthetically in future power systems. In depth knowledge about the amount of inertia provided by power consumers is very important for a future application of units supplying synthetic inertia. It strongly promotes the technical efficiency and cost effective application. A blackout in the city of Flensburg allows for a detailed research on the inertia contribution from power consumers. Therefore, power consumer categories are introduced and the inertia contribution is calculated for each category. Overall, the inertia constant for different power consumers is in the range of 0.09 to 4.24 s if inertia constant calculations are based on the power demand. If inertia constant calculations are based on the apparent generator power, the load inertia constant is in the range of 0.01 to 0.19 s. -

Moment of Inertia
MOMENT OF INERTIA The moment of inertia, also known as the mass moment of inertia, angular mass or rotational inertia, of a rigid body is a quantity that determines the torque needed for a desired angular acceleration about a rotational axis; similar to how mass determines the force needed for a desired acceleration. It depends on the body's mass distribution and the axis chosen, with larger moments requiring more torque to change the body's rotation rate. It is an extensive (additive) property: for a point mass the moment of inertia is simply the mass times the square of the perpendicular distance to the rotation axis. The moment of inertia of a rigid composite system is the sum of the moments of inertia of its component subsystems (all taken about the same axis). Its simplest definition is the second moment of mass with respect to distance from an axis. For bodies constrained to rotate in a plane, only their moment of inertia about an axis perpendicular to the plane, a scalar value, and matters. For bodies free to rotate in three dimensions, their moments can be described by a symmetric 3 × 3 matrix, with a set of mutually perpendicular principal axes for which this matrix is diagonal and torques around the axes act independently of each other. When a body is free to rotate around an axis, torque must be applied to change its angular momentum. The amount of torque needed to cause any given angular acceleration (the rate of change in angular velocity) is proportional to the moment of inertia of the body.