Dormancy and Growth Vigour of Seed Potatoes
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

LEGAL NOTICES WETLIKE KENNISGEWINGS 2 No
. March Vol. 657 Pretoria, 27 M t 2020 aar No. 43149 ( PART1 OF 2 ) LEGAL NOTICES WETLIKE KENNISGEWINGS 2 No. 43149 GOVERNMENT GAZETTE, 27 MARCH 2020 STAATSKOERANT, 27 MAART 2020 No. 43149 3 Table of Contents LEGAL NOTICES BUSINESS NOTICES • BESIGHEIDSKENNISGEWINGS Gauteng ....................................................................................................................................... 15 Eastern Cape / Oos-Kaap ................................................................................................................. 15 KwaZulu-Natal ................................................................................................................................ 16 Western Cape / Wes-Kaap ................................................................................................................ 16 COMPANY NOTICES • MAATSKAPPYKENNISGEWINGS Gauteng ....................................................................................................................................... 17 Western Cape / Wes-Kaap ................................................................................................................ 17 LIQUIDATOR’S AND OTHER APPOINTEES’ NOTICES LIKWIDATEURS EN ANDER AANGESTELDES SE KENNISGEWINGS Gauteng ....................................................................................................................................... 20 Limpopo ....................................................................................................................................... 20 Western -
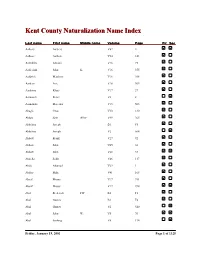
Kent County Naturalization Name Index, Aalbers, A
Kent County Naturalization Name Index Last name First name Middle name Volume Page Fir Sec Aalbers Aalbers V62 4 Aalbers Aalbert V24 141 Aalddriks Antonie V16 75 Aalderink John K. V16 355 Aaldrick Matthew V16 308 Aardem Arie V16 304 Aardema Klaas V17 27 Aarnouds Pieter V6 8 Aarnoudse Marenus V15 503 Abagis Chas V30 130 Abbas Sain Allez V49 265 Abbelma Joseph B1 F5 Abbelma Joseph V2 564 Abbott Frank V27 92 Abbott John V45 36 Abbott John V68 33 Abdella Salik V46 117 Abdo Ahamad V29 1 Abdoo Mike V41 168 Abeaf Moses V17 391 Abeaf Moses V17 394 Abel Frederick FW B1 F1 Abel Gustav B1 F4 Abel Gustav V2 540 Abel John W. V5 70 Abel Ludwig V8 134 Friday, January 19, 2001 Page 1 of 1325 Last name First name Middle name Volume Page Fir Sec Abella Salih V68 85 Abezi Albert V25 76 Aboabsee Theab V74 40 Aboasee Theab V18 150 Abood N. B1 F5 Abood Nemy V3 90 Abraham John V17 381 Abrahamson Charles Y. B7 200 Abram John B1 F1 Abramson Morris B1 F3 Abraursz Abram Peter V27 159 Abromaitis Louis V27 381 Abromaitis Louis V67 90 Absmaier Carl V77 4 Accardi Guiseppe V50 79 Acheson John V16 616 Achille Minciotti V51 142 Achtenhof Jakob V15 145 Achter Jan V17 200 Achterhof Henri B1 F1 Achterhof Johannes V15 500 Achterhof Matheus B1 F1 Achtjes John B7 107 Ackermann Joseph V15 282 Acton John C. B7 222 Adair David G. V15 335 Adair Joseph V15 335 Adalphson Emil V18 197 Friday, January 19, 2001 Page 2 of 1325 Last name First name Middle name Volume Page Fir Sec Adam Frickartz Heinrich V41 279 Adama Jelle V22 176 Adamawiczus Baltris V37 155 Adamczak Peter V38 245 Adamczyk Wladyslaw V35 291 Adams Edward John V24 70 Adams Frank B1 F2 Adams George W. -

Correspondence of Maria Van Rensselaer (1669-1689)
CORRESPONDENCE OF MARIA VAN RENSSELAER 1669-1689 Translated and edited by A. J. F. VAN LAER Archivist, Archives and History Division ALBANY THE UNIVERSITY OF THE STATE OF NEW YORK I 935 PREFACE In the preface to the Correspondence of Jeremias van Rens selaer, which was piiblished in 1932, attention was called to the fact that after the death of Jeremias van Rensselaer his widow carried on a regular correspondence with her husband's youngest brother, Richard van Rensselaer, in regard to the administration of the colony of Rensselaerswyck, and the plan was announced to publish this correspondence in another volume. This plan has been carried into effect in the present volume, which contains translations of all that has been preserved of the correspondence of Maria van Rensselaer, including besides the correspondence with her brother-in-law many letters which passed between her and her brother Stephanus van Cortlandt and other members of the Van Cortlandt family. Maria van Rensselaer was born at New York on July 20, 1645, and was the third child of Oloff Stevensen van Cortlandt and his wife Anna Loockermans. She married on July 12, 1662, when not quite 17 years of age, Jeremias van Rensselaer, who in 1658 had succeeded his brother Jan Baptist van Rensse laer as director of the colony of Rensselaerswyck. By him she had four sons and two daughters, her youngest son, Jeremias, being born shortly after her husband's death, which occurred on October 12, 1674. As at the time there was no one available who could succeed Jeremias van Rensselaer as director of the colony, the burden of its administration fell temporarily upon his widow, who in this emergency sought the advice of her brother Stephanus van Cortlandt. -

Show #B0612 (Tonya Brown): 2018 Stars and Stripes (06/29/2018-07/01/2018) 1
© An Equine Production Released to The Equine Chronicle with permission. Sponsored by Shadow Trailers Show #B0612 (Tonya Brown): 2018 Stars and Stripes (06/29/2018-07/01/2018) 1. AQHA 212002: L1 Amt Showmanship at Halter - Shown: 19 PlaceBack# Horse's Name Rider's Name Owner's Name Owner's City 1 15 Sleepin In The Cafe Rowdy Bentham Rowdy Bentham Appleton, WI 2 125 Good As Expected Kaylee Swanigan Angela Mccordic Stanhope Wadmalaw IS , SC 3 108 A Good Luck Kiss Janis Simmons Janis Simmons Navasota, TX 4 131 Made With Real Cocoa Daniel J Maletic Daniel J Maletic Annapolis, MD 5 35 Surely I Will Brittany Pritchett Jenny Pritchett Marietta, GA 6 105 Sumdiscretionadvised Liz Harris Liz Harris Melbourne, FL 7 204 Shady Connection Sarah J O'Hara Sarah J O'Hara Huntsville, AL 8 155 Irishironsr Smilin Taylor Battistelli Taylor Battistelli Port St Lucie, FL 9 201 Hez Naturally Hot Susan Basquez Kristin Basquez Port Saint Lucie, FL 2. AQHA 212003: L2 Amt Showmanship at Halter - Shown: 13 PlaceBack# Horse's Name Rider's Name Owner's Name Owner's City 1 198 Invested In Cowboys Lisa K Mazurka Lisa K Mazurka Sutton, MA 2 91 Good Weekend Leah Burgess Leah Burgess Williamsport, PA 3 217 Good To Be Envied Farley Wickre Farley Wickre Kitty Hawk, NC 4 61 SH Counting Stars Kara M Flentje Kara M Flentje Verona, WI 5 153 Assets Of The Stars Megan Hunt Megan Hunt Byhalia, MS 6 822 Invited To A Goodbar Shannon M Thrower Shannon M Thrower Orrum, NC 7 35 Surely I Will Brittany Pritchett Jenny Pritchett Marietta, GA 8 34 IE Thumptin Special Darian Robin Sullivan Darian Robin Sullivan Hartsville, TN 9 89 Good At Da Bar Rebecca Fussell Rebecca Fussell Cedar Hill, TX 10 92 Dreamin So Good Molly Mckechnie Molly Mckechnie Topsham, ME 11 247 Dun Ben Noticed Shelley Dawn Dr Grissom Shelley Dawn Dr Grissom Aubrey, TX 12 155 Irishironsr Smilin Taylor Battistelli Taylor Battistelli Port St Lucie, FL 3. -

Review of Web Mapping: Eras, Trends and Directions
International Journal of Geo-Information Review Review of Web Mapping: Eras, Trends and Directions Bert Veenendaal 1,*, Maria Antonia Brovelli 2 ID and Songnian Li 3 ID 1 Department of Spatial Sciences, Curtin University, GPO Box U1987, Perth 6845, Australia 2 Department of Civil and Environmental Engineering (DICA), Politecnico di Milano, P.zza Leonardo da Vinci 32, 20133 Milan, Italy; [email protected] 3 Department of Civil Engineering, Ryerson University, 350 Victoria Street, Toronto, ON M5B 2K3, Canada; [email protected] * Correspondence: [email protected]; Tel.: +618-9266-7701 Received: 28 July 2017; Accepted: 16 October 2017; Published: 21 October 2017 Abstract: Web mapping and the use of geospatial information online have evolved rapidly over the past few decades. Almost everyone in the world uses mapping information, whether or not one realizes it. Almost every mobile phone now has location services and every event and object on the earth has a location. The use of this geospatial location data has expanded rapidly, thanks to the development of the Internet. Huge volumes of geospatial data are available and daily being captured online, and are used in web applications and maps for viewing, analysis, modeling and simulation. This paper reviews the developments of web mapping from the first static online map images to the current highly interactive, multi-sourced web mapping services that have been increasingly moved to cloud computing platforms. The whole environment of web mapping captures the integration and interaction between three components found online, namely, geospatial information, people and functionality. In this paper, the trends and interactions among these components are identified and reviewed in relation to the technology developments. -

V.V. Bavel 1 TOTO
V.V. Bavel 1 TOTO Stand Deelnemer Totaalscore Stand Deelnemer Totaalscore 1 Robbert Matthijs 26 50 Marian van Gils 15 2 Mark de Vroom 26 51 Henrie van Gils 15 3 Robbert Matthijs 26 52 Luuk van den Berg 15 4 Bart van Beek 23 53 Ray van den Berg 15 5 Sven van der Maaten 23 54 Ralph van Loon 15 6 Pieterbas Plasman 23 55 Frans Oomen 15 7 Paul Carton 23 56 Daan Kriellaars 15 8 Shane Lourens 21 57 Caroline Bauwens 15 9 Mike Polderman 21 58 Bouke Kramer 15 10 Mery Peters 21 59 Adrienne Polderman 15 11 Kees Schaerlaeckens 21 60 Ruud Franken 15 12 Jeffry Kools 21 61 Paul Adriaansen 15 13 Fabienne Peters 21 62 Yvette de Vroom 15 14 Cees Adriaansen 21 63 Lucien Diepstraten 15 15 Anita van Loon 21 64 JP Huijskens 15 16 Lucy de Wijs 21 65 Jacobien Dekkers 15 17 Kim Leijten 21 66 Esther Plasman 15 18 Stijn Adriaansen 21 67 Daan Diepstraten 15 19 Aisha Valk 20 68 Corne Peeters 15 20 Valentijn Koomans 20 69 Bas Plasman SR 15 21 Colinda van den Maagdenberg 20 70 Ad van Gorkum 15 22 Toine Carton 20 71 Werner de Heer 12 23 Rik van Oosterwijck 20 72 Wout Dekkers 12 24 Jan Bossers 20 73 Corne vd Kieboom 12 25 Corie Oomen 20 74 Robbert Matthijs 12 26 JP Huijskens 20 75 Eva Carton 12 27 Stef Horsten 18 76 JP Huijskens 12 28 Tessa Donkers 18 77 Martijn Willemsen 12 29 Niek Akkermans 18 78 Teun Adriaansen 12 30 Julie van Noesel 18 79 Sander van Gils 12 31 Dirk van Dijk 18 80 Maarten van Loon 12 32 Wim Meeuwisse 18 81 Jeroen Theunis 12 33 Pim Schaerlaeckens 18 82 Jack Jansen 12 34 Rick van den Maagdenberg 18 83 Hans Joppen 12 35 JP Huijskens 18 84 Mike Marijnissen 12 36 Job Horsten 18 85 Giel Dekkers 12 37 Bart Diepstraten 18 86 Giel Carton 11 38 Will de Vroom 18 87 Marco Verschuren 9 39 Teun van den Kieboom 18 88 Thomas Adolphs 9 40 Roeland Verbeet 18 89 Ilse Geers 9 41 JP Huijskens 18 90 Rob Kriellaars 6 42 Paul Kriellaars 18 91 Peter Dekkers 6 43 Ruud van den Maagdenberg 18 44 Inge Koomans 18 45 Robbert Matthijs 17 46 Christel Geerts 17 47 Rob Roelands 17 48 Gijs Pheninckx 17 49 Jurgen van Loon 15. -

Omslag Gekleurd Papier
Burgumers, verhalen zelf Uitgave Nut Bergum Burgumers, verhalen zelf Een verhalen bundel voor en door (oud) Burgumers Uitgave Nut Bergum e.o. – Redactie Taeke Doevendans 2011 1 2 Voorwoord Een boek geschreven door en voor Burgumers (en daarbuiten), onder de titel “Burgumers, verhalen zelf”. Een vervolg op de vraag van het Nut Bergum aan mij om over oud plaatsgenoten te schrijven. Een megaklus waarop ik niet ben ingegaan. Wel het idee sprak me aan, omdat er een generatie vertellers en schrijvers over “gewone” recente dorpshistorie verdwijnt. Hetzelfde leefde bij mij reeds als fotograaf rond 1975 toen ik een fotoboek maakte over “Wâldman” Douwe Postma. Deze fotoserie is dan ook de start geweest als tentoonstelling in de bibliotheek en “Berchhiem”. Via een verzoek aan de Nutleden en de media is daarna gevraagd een bijdrage te schrijven over Burgumers en hun belevenissen, beroepen en anekdotes. Het is geworden wat u hierin kunt lezen, een mooie bloemlezing over uw dorp en haar inwoners. Naast aanvullingen kwam het vooral tot stand door de inzet van de inzenders Dirk Scherjon, Rinze van der Wal, Jeltsje Haisma-Boskma, Pieter Bekkema, Klaas Henstra, Jittie Bosma-Hoekema, Gelske Hoekstra-Postma, opmaak Wichard Dijkstra en stijl- en taalcorrector Fokke Broers. Ik ben blij dat het Nut hiermee kan laten zien dat locale betrokkenheid kan leiden tot een “eigen” uitgave. Over wat zich hier ter plaatse heeft afgespeeld, beschreven letterlijk uit de eerste hand. Hopelijk geniet u er net zo van als ik. Veel leesplezier in herkenning en verbazing. Taeke Doevendans / voorzitter Nut en redacteur 3 4 Inhoudsopgave Voorwoord 3 Inhoudsopgave 5 Historie Nut departement Bergum e.o. -

Township Public Schools
Regular Meeting Minutes - May 3, 2018 Page 1 of 220 Township Pub lic Schools REGULAR MEETING MINUTES MAY 3,2018 The Regular Meeting of the Wayne Township Board of Education was held on Thursday, May 3, 2018 in Council Chambers of the Municipal Building at 475 Valley Road, Wayne, New Jersey 07470. Executive Session was held in the Large Conference Room, in the Administration Building, 50 Nellis Drive, Wayne, New Jersey 07470. The meeting was scheduled to begin at 6:00 p.m. , pursuant to the terms of the Sunshine Law. The meeting was called to order at 6:05 p.m. by Donald Pavlak, Jr. , Board President. I. Reading of Sunshine Law statement Adequate notice of this Regular and Executive Meeting, setting forth time, date and location, has been provided in accordance with the requirements of the Open Public Meetings Act on January 9, 2018 by: Prominently posting a copy on the bulletin board in the lobby of the offices of the Board of Education, which is a public place reserved for such announcements, transmitting a copy of this notice to The Record, The Wayne Today, and the Municipal Clerk. II. Roll Call PRESENT: Eileen Albanese, Mitch Badiner, Michael Bubba, Cathy Kazan, Gail Okun, Donald Pavlak, Jr., Suzanne Pudup, and Stacey Scher. ABSENT: Sean Duffy ALSO PRESENT: Dr. Mark Toback, Superintendent, Donna Reichman, Assistant Superintendent, Jinnee DeMarco, Assistant School Business Administrator/Board Secretary and John Geppert, Board General Counsel. Amotion was made to convene into Executive Session. Regular Meeting Minutes - May 3, 2018 Page 2 of 220 III. Approval to Convene into Executive Session X-2018-1 RECOMMENDED ACTION: Approved WHEREAS, the Wayne Township Board of Education (hereinafter referred to as the "Board") from time to time must convene into Executive Session to discuss confidential matters including but not limited to personnel issues, legal matters, student issues and labor negotiations. -

Genealogy of the Doremus Family in America
GENEALOGY — —OF THE DOREMUS FAMILY IN AMERICA: Descendants of Coknelis Doremus, from Breskens and mlddleburg, in holland, who emigrated to amer- ica about 1685-6,and shtl'i.kt) at acquacka- nonk (now Paterson), New Jersey. WILLIAMNELSON. PATERSON, N.L: THE PRESS PRINTING AND PUBLISHING COMPANY. 1897. Doremus Genealogy ifzA-y y-ViAAx) x GENEALOGY — — OF THE DOREMUS FAMILY IN AMERICA : Descendants of Cornelis. Doremus, from Breskens and . mlddelburg, in holland, whoemigrated to amer- ica about 1685-6, and settled at acquacka- nonk (now Paterson), New Jkrsey. WILLIAM NELSON. PATERSON, N. J. : TTIE PRESS PRINTING AND PUBLISHING COMPANY. c^1 y < One Hundred, Copies Printed. / & <>v TO MY WIFE SALOME WILLIAMSDOREMUS NELSON THIS RECORD OFHER ANCESTRY' AND KINSFOLK IS DEDICATED. FOREWORDS. Gj'HE material for the history of the Doremus Family for y;the first four or five generations, as given in the follow- oat}\i*Tf»A ' mo r»acr#»c Vine ViA#»n liv frit**writer nlmnct *»V- ing pages, has been gathered by the writer almost ex- clusively from original investigations in church registers, records of deeds and wills, and from tombstones. For the accounts of the later generations, and particularly of those scattered far from their ancestral homes, he has had to rely largely upon correspondence, often dilatory and otherwise unsatisfactory, but in very many cases prompt, intelligent and interested. It would be a pleasure, did not delicacy forbid, to mention some of those who have cordially re- sponded to the author's requests for information, and who have thus materially aided inmaking this little work more complete. -

Creation and Christ
Wissenschaftliche Untersuchungen zum Neuen Testament · 2. Reihe Herausgeber / Editor Jörg Frey (Zürich) Mitherausgeber / Associate Editors Markus Bockmuehl (Oxford) · James A. Kelhoffer (Uppsala) Tobias Nicklas (Regensburg) · Janet Spittler (Charlottesville, VA) J. Ross Wagner (Durham, NC) 527 Angela Costley Creation and Christ An Exploration of the Topic of Creation in the Epistle to the Hebrews Mohr Siebeck Angela Costley, born 1984; 2005 BA Hons in Theology, 2008 PG Cert in the Theory of Education from Durham University; 2011 MSt in Jewish Studies from the University of Oxford; 2018 PhD from St. Patrick’s College, the Pontifical University, Maynooth. orcid.org / 0000‑0001‑8000‑7277 ISBN 978‑3‑16‑156502‑1 / eISBN 978‑3‑16‑156530‑4 DOI 10.1628 / 978‑3‑16‑156530‑4 ISSN 0340‑9570 / eISSN 2568‑7484 (Wissenschaftliche Untersuchungen zum Neuen Testament, 2. Reihe) The Deutsche Nationalbibliothek lists this publication in the Deutsche Nationalbiblio‑ graphie; detailed bibliographic data are available at http://dnb.dnb.de. © 2020 Mohr Siebeck Tübingen, Germany. www.mohrsiebeck.com This book may not be reproduced, in whole or in part, in any form (beyond that per‑ mitted by copyright law) without the publisher’s written permission. This applies partic‑ ularly to reproductions, translations and storage and processing in electronic systems. The book was typeset and printed on non‑aging paper by Laupp & Göbel in Gomaringen, and bound by Buchbinderei Nädele in Nehren. Printed in Germany. For my parents, without whose support this thesis would not have been possible. Preface This work is a revised version of my PhD thesis, completed at St. Patrick’s Col‑ lege, the Pontifical University, Maynooth between 2013 and 2018, under the supervision of Rev. -

0.00 7/8/18 GQHA Stars & Stripes, Big a and Sh
Show Name : GQHA Stars & Stripes, Big A and Show For Dough AQHA 2 (2018) 7/8/18 Class Name : Open Ranch Riding Class Entries : 0 Money Awarded : $0.00 Place Horse Name : Exhibitor Name : Owner Name: Money Earned Show Name : GQHA Stars & Stripes, Big A and Show For Dough AQHA 2 (2018) 7/8/18 Class Name : Amateur Ranch Riding Class Entries : 0 Money Awarded : $0.00 Place Horse Name : Exhibitor Name : Owner Name: Money Earned Show Name : GQHA Stars & Stripes, Big A and Show For Dough AQHA 2 (2018) 7/8/18 Class Name : Novice Amateur Ranch Riding Class Entries : 0 Money Awarded : $0.00 Place Horse Name : Exhibitor Name : Owner Name: Money Earned Show Name : GQHA Stars & Stripes, Big A and Show For Dough AQHA 2 (2018) 7/8/18 Class Name : Youth Ranch Riding Class Entries : 0 Money Awarded : $0.00 Place Horse Name : Exhibitor Name : Owner Name: Money Earned Show Name : GQHA Stars & Stripes, Big A and Show For Dough AQHA 2 (2018) 7/8/18 Class Name : Novice Youth Ranch Riding Class Entries : 0 Money Awarded : $0.00 Place Horse Name : Exhibitor Name : Owner Name: Money Earned © National Snaffle Bit Association www.nsba.com Show Name : GQHA Stars & Stripes, Big A and Show For Dough AQHA 2 (2018) 7/8/18 Class Name : Green Western Pleasure Class Entries : 9 Money Awarded : $0.00 Place Horse Name : Exhibitor Name : Owner Name: Money Earned 1 Made My Way Cody Parrish Barbara R Cliford $0.00 2 Zip N Machine Bret Parrish Adrienne Evans $0.00 3 Two Ziplines Taft Dickerson Katie Kopf $0.00 4 Million To 1 Jay Starnes Nettie Olsen $0.00 5 An Absolute Martini -

The Ancestry and Posterity of Cornelius Henry Tiebout of Brooklyn
W-i tf 5: I "7 Di 558 1 O"i 'If ¦«**? [;< s= '•*'^««, r f S7 JS? t! &5i IHS^{ mfmst! ;*? C ' -^sia'iaffj?Ja ! k*^ :'?sM£ •lafe.^ i*t 5 j£^??f a'f » *s :"^Si' i 1 tit |«. f?a?J #:-< E^ E^ E^* E^* In E** Em E^* 14: E»* =*** ~ __— i <i =» ? a 32 Mg < o 5 ' ' =«* g s § E* 8 S =IA E^ i« = M =—• THE ANCESTRY AND POSTERITY OF CORNELIUS HENRY TIEBOUT OF BROOKLYN Copy No. **/, PRESENTED TO Jfr£&&€££</ /^^.^L / With the Compliments of PRINTED FOR PRIVATE DISTRIBUTION 1 \* *%* Itis a pleasure to state as a foreword that itis solely because of the interest in and knowledge of matters genea logical of Mr.Francis V.Morrell, a son of one of my former employers, that this work has been produced. To him we are indebted for most all of the documentary information, the arrangement of the book and its complete index, as well as the careful editing of fragmentary family records and the solution of perplexing problems of descent. 2 TIEBOUT This name is Teutonic, appearing as Tybalt, meaning people's prince. Itis found in several forms, among them the Danish, Theobald; Dutch, Tiebout; French, Thibaut; German, Dietbold, and Italian, Teobaldo. It has also appeared Thieboult. STANDARD DICTIONARY. The following spellings are found inAmerican records : Tybout, Tibout, Tibouwt, Thibou, Tiebout, Tiebouwt, Tiebaut, Thibo, Teabout, Thebout, Thebaut, Thibout, Tebow, Tibouts, Tiboe, Tebou, Tebout, Tebow, Tebo, Thibe, Tibau and Thiboutszen, plural or possessive. The name was known in Flanders when the whole country was under French rule and the principal seat of the family was at Bruges; from thence the American branch sprung.