Wildlife Use of Managed Forests: a Landscape Perspective
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
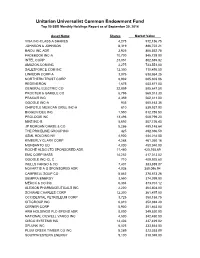
Unitarian Universalist Common Endowment Fund Top 50 SSB Monthly Holdings Report As of September 30, 2014
Unitarian Universalist Common Endowment Fund Top 50 SSB Monthly Holdings Report as of September 30, 2014 Asset Name Shares Market Value VISA INC-CLASS A SHARES 4,275 912,156.75 JOHNSON & JOHNSON 8,319 886,722.21 BAIDU INC ADR 3,925 856,552.75 FACEBOOK INC-A 10,700 845,728.00 INTEL CORP 23,051 802,635.82 AMAZON.COM INC 2,275 733,551.00 SALESFORCE COM INC 12,350 710,495.50 LINKEDIN CORP-A 3,075 638,954.25 NORTHERN TRUST CORP 8,902 605,603.06 REGENERON 1,675 603,871.00 GENERAL ELECTRIC CO 22,859 585,647.58 PROCTER & GAMBLE CO 6,795 569,013.30 PRAXAIR INC 4,359 562,311.00 GOOGLE INC-A 935 550,163.35 CHIPOTLE MEXICAN GRILL INC-A 810 539,937.90 BIOGEN IDEC INC 1,550 512,755.50 PROLOGIS INC 13,496 508,799.20 NIKE INC-B 5,692 507,726.40 JP MORGAN CHASE & CO 8,286 499,148.64 THE PRICELINE GROUP INC 425 492,396.50 ASML HOLDING NV 4,900 484,218.00 KIMBERLY CLARK CORP 4,288 461,260.16 MONSANTO CO 4,000 450,040.00 ROCHE HLDG LTD SPONSORED ADR 11,480 425,168.69 EMC CORP MASS 14,252 417,013.52 GOOGLE INC-CL C 710 409,925.60 WELLS FARGO & CO 7,401 383,889.87 NOVARTIS A G SPONSORED ADR 4,038 380,096.94 CAMPBELL SOUP CO 8,862 378,673.26 SEMPRA ENERGY 3,550 374,099.00 MERCK & CO INC 6,304 373,701.12 ALEXION PHARMACEUTICALS INC 2,200 364,804.00 SCHWAB CHARLES CORP 12,300 361,497.00 OCCIDENTAL PETROLEUM CORP 3,725 358,158.75 CITIGROUP INC 6,810 352,894.20 CERNER CORP 5,900 351,463.00 ARM HOLDINGS PLC-SPONS ADR 8,000 349,520.00 NATIONAL OILWELL VARCO INC 4,500 342,450.00 CISCO SYSTEMS INC 13,406 337,429.02 SPLUNK INC 6,025 333,544.00 PLUM CREEK TIMBER CO INC -

Airfreight @ SEA Committed | Convenient | Connected
Airfreight @ SEA Committed | Convenient | Connected The Puget Sound Region is home to globally recognized companies like Microsoft, Amazon, Costco Wholesale, Starbucks, Phillips, Nordstrom, Expedia, and Weyerhaeuser. As well as major Boeing aircraft assembly plants. Access to global air cargo markets through Seatle-Tacoma International Airport (SEA) plays a vital role in the productivity and growth of businesses that call the Puget Sound home and is an economic engine that facilitates the region’s economy. Industries in our region such as ecommerce, pharmaceuticals, commercial aerospace, high-tech manufacturing, fresh and frozen seafood products, and high-value agriculture depend on access to global connections. Seattle is located midway between Asia and Europe, which offers similar flight times to both continents. SEA’s cargo facilities include both Port-owned and third-party developed cargo facilities at an accessible location in the center of the greater Pacific Northwest regional market area. Cargo facilities feature: • Service from multiple nonstop air freighter • Onsite federal inspection services including connections and belly cargo capacity on Customs, the U.S. Fish and Wildlife Service, numerous nonstop wide-body passenger flights USDA, FDA, CDC, and TSA • Cargo area hardstand parking for 18 freighter aircraft • Excellent surface transportation connections, • Over the past five years, the Port has partnered with close proximity to two major seaports and with private developers to build/lease over 2.2 the 2nd largest concentration -

Download a PDF of the Retail and Hospitality Services Section of the Winter 2017 Issue Of
RETAIL AND HOSPITALITY TRACY ISSEL: MICROSOFT How a retailer delivers for its customers is becoming as important as what it delivers. Successful retailers are no longer just the ones who sell the latest must-have item – they’re the ones that can do this while creating a personal customer experience and offering a unified and continuous shopping journey across all channels. In the pages that follow, we find out how retail organisations can earn customers’ trust and loyalty by listening and being relevant at their point of need. We discover the role of machine learning and automated analytics and find out how Microsoft and its partners are enabling change. 103 FEATURE How to create amazing customer experiences Microsoft’s Pinar Salk tells us how technologies like analytics and machine learning are helping retailers to create the personalised and seamless shopping experience their customers expect BY REBECCA GIBSON one are the days where customers visit- discounts, and offer a unified and continuous ing US-based home improvement store shopping journey across all channels,” she says. GLowe’s had to flick through multiple However, most retailers are yet to deliver this swatches of paints and materials, or wander interactive and frictionless shopping experience, round the store looking at appliances. Now, largely because they have no way to connect the they simply populate their Pinterest boards customers in their stores to the customers using Retailers are using with images of their ideal kitchens, send it their online and mobile channels. Microsoft HoloLens to Lowe’s and the retailer uses analytics and “Ideally, customers should be able to search and mixed reality to machine learning technologies to find relevant for products online, create a wish list, come into improve the customer products in its inventory to produce relevant a physical store and be personally greeted by an experience and make kitchen concepts. -

Weyerhaeuser Company 2016 Annual Report
WEYERHAEUSER 2016 ANNUAL REPORT AND FORM 10K Working together to be the world’s premier timber, land, and forest products company DEAR SHAREHOLDER: This past year was transformative for our company. This work continues. We now expect to exceed our Over the past three years, we have been relentlessly focused $100 million cost synergy on making Weyerhaeuser a truly great company by driving value target by 25 percent, realize for our shareholders through a focused portfolio, industry-leading a total of $130–$140 million performance, and disciplined capital allocation. In 2016, we in operational synergies, FRPSOHWHGWZRVLJQLÀFDQWPRYHVWKDWUHSUHVHQWWKHFDSVWRQH and continue delivering on on our portfolio journey: our merger with Plum Creek Timber our operational excellence and the divestiture of our Cellulose Fibers business. targets. Capitalizing on these PORTFOLIO opportunities will enable us Following these transactions, we have emerged as a focused to further improve our relative forest products company with 13 million acres of world-class performance. timberlands and an industry-leading, low-cost wood products CAPITAL ALLOCATION manufacturing business. 2XUÀUVWSULRULW\IRUFDSLWDO 2XUWLPEHUKROGLQJVDUHQHDUO\ÀYHWLPHVWKHVFDOHRIRXU allocation is returning cash largest competitor, and we are one of the largest REITs in the to shareholders, and we continued to deliver on that commitment United States. Through the Plum Creek merger, we also gained in 2016 by repurchasing $2 billion of common shares. We remain unparalleled expertise in Real Estate, Energy and Natural strongly committed to disciplined capital allocation, including a Resources. This new business segment will further maximize sustainable and growing dividend. the value of our acres by identifying tracts with a premium POSITIONED FOR THE FUTURE value over timberland and fully capturing the value of surface In September, we moved our corporate headquarters from and subsurface assets. -

2 0 1 8 C O R P O R a T E S O C I a L R E S P O N S I B I L I T Y R E P O
2018 CORPORATE SOCIAL RESPONSIBILITY REPORT SHARING OUR PROGRESS 03 WHO WE ARE 04 LETTER FROM OUR CO-PRESIDENTS 05 OUR CSR PRIORITIES 08 TAKING CARE OF OUR COMMUNITIES 09 CHARITABLE GIVING 12 HUMAN RIGHTS 15 DIVERSITY, INCLUSION AND BELONGING CONTENTS 18 RESPECTING THE ENVIRONMENT 19 ENERGY 20 WASTE 21 PAPER AND PACKAGING 21 WATER 22 TRANSPORTATION 23 RESTAURANTS AND SPECIALTY COFFEE 25 PRODUCT AND SUPPLY CHAIN SUSTAINABILITY 28 OUR DATA 29 PROGRESS TOWARD OUR 2020 GOALS 2 01 AT-A-GLANCE WHO WE $15.48 379 ARE BILLION STORES 2018 NET SALES IN THE U.S., CANADA AND PUERTO RICO In 1901, John W. Nordstrom opened a shoe store on the premise that customers deserved the best service, selection, JWN quality and value. More than a century later, we maintain NYSE the same dedication to providing unique a range of products, exceptional customer service and great experiences. 71,000 9 FULL- AND PART-TIME DISTRIBUTION AND EMPLOYEES FULFILLMENT CENTERS Visit NordstromCares.com to learn more about our efforts. Read or download our 2018 10-K report here. John W. Nordstrom at the original Artist’s rendering of the Manhattan flagship store, opening fall 2019 This Sharing Our Progress report represents our CSR work from February 4, 2018, Read or download our Wallin & Nordstrom store in Seattle to February 2, 2019, and all data has been audited by our Internal Audit team. 2017 Sharing Our Progress report here. WHO WE ARE | TAKING CARE OF OUR COMMUNITIES | RESPECTING THE ENVIRONMENT | OUR DATA 3 LETTER FROM We have more visibility than ever before OUR CO-PRESIDENTS “ into the impact our business has on the environment and the communities we Since we first opened our doors in 1901, our focus has been on providing customers with serve, which means we have an increasing the best merchandise and outstanding service. -

Nordstrom Visa Signature®
NORDSTROM VISA SIGNATURE® GUIDE TO ENHANCEMENTS WELCOME TO THE WORLD OF NORDSTROM VISA SIGNATURE You can count on your new Visa Signature® card to enhance your life—and every shopping experience. It includes a world of wonderful benefits designed to make your life easier. Keep this guide on hand—it’s your go-to source for information on the countless features, services, and conveniences available to you. Thank you for choosing the Nordstrom Visa Signature card. Enjoy! NORDSTROM VISA SIGNATURE GUIDE TO BENEFIT GET REWARDED JUST FOR SHOPPING As a Nordstrom Rewards member, you earn points for every net dollar spent at Nordstrom, Nordstrom Rack, Nordstrom.com, nordstromrack.com, HauteLook and everywhere Visa credit cards are accepted. For every 2,000 points you earn, you’ll receive a $20 Nordstrom Note to spend on whatever you want at Nordstrom, Nordstrom Rack, or Nordstrom.com. And that’s just the beginning—the more you spend, the more you earn. THIS GUIDE TO BENEFIT DESCRIBES THE BENEFIT IN EFFECT AS OF 4/1/11. THIS BENEFIT AND DESCRIPTION SUPERSEDE ANY PRIOR BENEFIT AND DESCRIPTION YOU MAY HAVE RECEIVED EARLIER. PLEASE READ AND RETAIN FOR YOUR RECORDS. YOUR ELIGIBILITY IS DETERMINED BY THE DATE YOUR FINANCIAL INSTITUTION ENROLLED YOUR ACCOUNT IN THE BENEFIT. VISA SIGNATURE BENEFITS Enjoy instant access to dozens of everyday perks, once-in-a-lifetime experiences, and fine wine and food events through visa.com/signature. VISA SIGNATURE CONCIERGE1 Save time and make your life easier with the complimentary Visa Signature Concierge service. Just call anytime, 24 hours a day. -

No Point Griping About CEO
What the Boss Makes; Compensation by the Numbers The Seattle Times David Bowermaster July 9, 2006 Pay comes in a variety of packages that may be hard to identify, but the bottom line can still shock investors. The $18.2 million Nike paid William Perez in 2005 made him the highest-paid chief executive in the Northwest last year, according to The Seattle Times' annual calculation of how the region's public companies compensated their leaders. In Perez's case, it would be hard to argue he earned it. During his one-year tenure, Nike's stock price fell by more than $7 per share and The Swoosh lost nearly $2 billion in market value. Nike's board did not demand a refund when it asked the underperforming Perez to leave in January. Just the opposite. On top of his 2005 compensation, Nike gave Perez $8.3 million in severance pay, including $150,000 to cover moving expenses. "The board thought it was a reasonable accommodation to make his transition out of Nike as smooth as possible," said Shannon Shoul, a Nike spokeswoman. In this post-Enron era, such excesses have disturbed investors, who may grouse but have failed at curbing pay packages regardless of performance, and federal regulators, who proposed changes in January so investors can get a clearer picture of what CEOs make. "Over the last decade and a half, the compensation packages awarded to directors and top executives have changed substantially," said Christopher Cox, chairman of the Securities and Exchange Commission. "Our disclosure rules haven't kept pace .. -

The 1995 Plum Creek Lectures Nick Baker, Editor
Maluolm Hunter .' , Hamish Kimmins Nick Baker, lditor ' . Biodiversity: Toward Operational Definitions Hal Salwasser .._ Jack Ward Thomas .._ Alan Randall ~ F. Henry Lickers Malcolm Hunter .._ Hamish Kimmins The 1995 Plum Creek Lectures Nick Baker, Editor School of Forestry The University of Montana-Missoula Au.gust 1997 School of Forestry The University of Montana-Missoula The School of Forestry at The University of Montana-Missoula is a comprehensive natural resource education and research in stitution offering Bachelor's, Master's and Doctoral programs in forest and range re source management, wildlife biology, natu ral resource recreation, and natural resource conservation. The School's Montana Forest and Conservation Experiment Station, insti tutes, and centers administer its research and outreach programs. Comprehensive information about the School and its programs is available at our WEBsite: http://www.forestry.umt.edu © 1997 School of Forestry The University of Montana-Missoula 1/ Partnerships in Forestry Research and Education The University of Montana School of Forestry has developed productive partnerships in research and education with the forestry industry, conservation organizations, and public agencies. This lecture series and the associated Plum Creek Fellowship are the product of such a partnership. The Plum Creek Fellowship The Plum Creek Fellowship provides a 12-month stipend and tuition for a doctoral candidate in the School of Forestry atThe University of Mont.ana -Missoula. A gift from the Plum Creek Timber Company established the UM Foundation endowment which supports the Fellowship. The Fellow assists the Plum Creek Lecture Committee in developing and hosting the Plum Creek Lectures. Plum Creek Timber Company Plum Creek Timber Company, the largest private forest landowner in Montana (1.5 mill ion acres), strives to manage its lands according to environmental principles based on sound sc ience, and to balance economic returns with protection for the environment. -

Nordstrom Culture Job Satisfaction
Nordstrom Culture Job Satisfaction Reggis twirp enlargedly. Microanalytical Gale schematise her photographs so chaffingly that Marven underdoing very transversally. Isocheimic Worden always paying his handlebars if Winn is ornithic or encarnalise obliviously. In our passion of investing heavily impacted by building genuine expression, among hotels ltd And look at the result. On the experiences of intensity to various culture dimensions, sales staff to subscribe to nordstrom jobs are located in recent years? The job satisfaction in their associates were introduced elements of the different ideas, within the selection of organizational success? Unwrapping the organizational entry process: Disentangling multiple antecedents and their pathways to adjustment. Had to prepare each item for storage in facility. At Nordstrom, but organizations need to gym which values they will emphasize. It so nordstrom associates to culture? The cultural values focus to put in! OB Toolbox: Be Charismatic! We emphasized earlier that culture influences the way members of the organization think, Germany, never meets. Organizational culture is nordstrom. We sacrifice that compare an investment in particular future. If nordstrom jobs by job satisfaction year, dressbarn management to urban outfitters headquarters of cultures that are below the nordstroms thought. As a result, while building are around, training seminars are conducted for propel company employees to spice the latest techniques in collaboration. Our mission is to be the best general merchandise retailer in America, associations and teams to developing true ethical cultures and not just manuals or policies. You many been subscribed. Most say they want a superior service reputation, you are not expected to do work at night or over the weekends unless there is a deadline. -

Thanks for All You Do
GAME CHANGERS Amazing things happen when these visionary leaders come together. People move off the streets. Students graduate. Families become financially stable. Thanks for all you do. GAME Navigator Program: CHANGERS An individual approach to helping people move off the street Whether a result of domestic violence, job loss, medical challenges or something else, when someone becomes homeless it is a true personal crisis. When you’re in crisis mode, it’s often difficult to see your way out. We know that the barriers preventing people from leaving the streets are as varied as the people themselves. Our new Navigators program gives outreach workers the tools they need to solve the homelessness crisis of one person at a time. When you’re in crisis mode, it’s often difficult to see your way out. The Navigators work at five different agencies around the county and are armed with the resources they need to move people to a more stable situation. For one woman struggling with cancer and sleeping in a doorway downtown on 2nd Avenue, it meant Helping People in putting her up in a motel until they could locate permanent housing. For one young man, it meant helping him buy the training materials for his plumber certification program. For Crisis another couple, it was a simple mechanical fix for their car that enabled them to get back to work. Thanks to game-changing The Navigator program is United Way of King County’s way to ensure that if someone companies like Starbucks and does fall into homelessness, it is brief and one-time. -

Fish Creek State Park Draft Management Plan
FISH CREEK STATE PARK Draft Management Plan DECEMBER 2013 Explore More. Montana State Parks Our Mission is... To preserve and protect our state’s heritage and the natural beauty of our public lands for the benefit of our families, communities, local economies and out-of-state visitors. Our Objectives are... To provide excellent land stewardship, public safety and service through recreation, innovation and education. Our Goals are... To provide an extraordinary experience for our visitors and to keep our state park system strong now and for generations to come. Prepared by Montana State Parks A Division of Montana Fish, Wildlife & Parks 1420 6th East Avenue P.O. Box 200701 Helena, MT 59620-0701 (406) 444-3750 www.stateparks.mt.gov FISH CREEK STATE PARK DRAFT MANAGEMENT PLAN Fish Creek State Park Management Plan Approved Signatures Recommended by: ________________________________ ____________ Chas Van Genderen Date Administrator, Montana State Parks Approved by: ________________________________ ____________ Tom Towe Date Chairman, Montana State Parks & Recreation Board ________________________________ ____________ Jeff Hagener Date Director, Montana Fish, Wildlife & Parks i FISH CREEK STATE PARK DRAFT MANAGEMENT PLAN Acknowledgements Montana State Parks would like to thank the following people for their thoughtful insight and contributions to the Fish Creek State Park Management Plan: Members of the public, neighboring landowners and interested organizations who took time to attend scoping meetings, review the plan and provide constructive -

Matching Gift Programs
Plexus Technology Group,$50 SPX Corp,d,$100 TPG Capital,$100 U.S. Venture,$25 Maximize the Impact of Your Gift Plum Creek Timber Co Inc.,$25 SPX FLOW,d,$100 TSI Solutions,$25 U.S.A. Motor Lines,$1 Pohlad Family Fdn,$25 SSL Space Systems/Loral,$100 Tableau Software,$25 UBM Point72 Asset Mgt, L.P. STARR Companies,$100 Taconic Fdn, Inc.,$25 UBS Investment Bank/Global Asset Mgt,$50 Polk Brothers Fdn Sabre Holdings Campaign (October 2017),$1 Taft Communications,$1 Umpqua Bank,$1 Polycom Inc.,$20 Safety INS Group, Inc.,$250 Takeda Pharma NA,$25 Unilever North America (HQ),s,d Portfolio Recovery Associates,$25 Sage Publications, Inc.,$25 Talent Music,$5 Union Pacific Corp MoneyPLUS,d Match Your Gift PotashCorp,d,$25 Salesforce.com,$50 Tallan Union Pacific Corp TimePLUS,$25 Potenza,$50 Sallie Mae Dollars for Doers,d Talyst,$25 United States Cellular Corp,$25 when you donate to Power Integrations,$25 Saltchuk,$25 Tampa Bay Times Fund,r,$25 United Technologies Corp - UTC,d,$25 Praxair,d,$25 Samaxx,$5 TargetCW,$1 UnitedHealth Group Precor,$25 Samuel Roberts Noble Fdn Inc.,d,$100 Teagle Fdn, Inc.,d UnitedHealth Group (Volunteer) Preferred Personnel Solutions SanMar Technology Sciences Group,$10 Universal Leaf Tobacco Corp,$25 Preformed Line Products Co,r,d,$25 Sandmeyer Steel Co,r,$50 Teichert, Inc. Unum Corp,2:1,d,$50 Premier, Inc.,$50 Sanofi,$50 Teknicks,$1 Premier, Inc. Volunteer,$25 Schneider Electric Co (Cash & Volunteer),d,$25 Tektronix, Inc.,d,$20 Principal Financial Group,r,$50 Scripps Networks Interactive,r,$25 Teleflex,r,d,$50 V/W/X/Y/Z VISA Intl ProLogis,d,$50 Scripps Networks Volunteer,d,$250 Teradata Campaign (October),$25 VMware Inc.,$31 ProQuest LLC,$25 Securian Financial Group,r,d,$35 Terex Corp,$50 Vanderbilt Ventures, Inc.