The Changing State of the Benguela Current Large
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

The Mean Flow Field of the Tropical Atlantic Ocean
Deep-Sea Research II 46 (1999) 279—303 The mean flow field of the tropical Atlantic Ocean Lothar Stramma*, Friedrich Schott Institut fu( r Meereskunde, an der Universita( t Kiel, Du( sternbrooker Weg 20, 24105 Kiel, Germany Received 26 August 1997; received in revised form 31 July 1998 Abstract The mean horizontal flow field of the tropical Atlantic Ocean is described between 20°N and 20°S from observations and literature results for three layers of the upper ocean, Tropical Surface Water, Central Water, and Antarctic Intermediate Water. Compared to the subtropical gyres the tropical circulation shows several zonal current and countercurrent bands of smaller meridional and vertical extent. The wind-driven Ekman layer in the upper tens of meters of the ocean masks at some places the flow structure of the Tropical Surface Water layer as is the case for the Angola Gyre in the eastern tropical South Atlantic. Although there are regions with a strong seasonal cycle of the Tropical Surface Water circulation, such as the North Equatorial Countercurrent, large regions of the tropics do not show a significant seasonal cycle. In the Central Water layer below, the eastward North and South Equatorial undercurrents appear imbedded in the westward-flowing South Equatorial Current. The Antarcic Intermediate Water layer contains several zonal current bands south of 3°N, but only weak flow exists north of 3°N. The sparse available data suggest that the Equatorial Intermediate Current as well as the Southern and Northern Intermediate Countercurrents extend zonally across the entire equatorial basin. Due to the convergence of northern and southern water masses, the western tropical Atlantic north of the equator is an important site for the mixture of water masses, but more work is needed to better understand the role of the various zonal under- and countercur- rents in cross-equatorial water mass transfer. -

Fronts in the World Ocean's Large Marine Ecosystems. ICES CM 2007
- 1 - This paper can be freely cited without prior reference to the authors International Council ICES CM 2007/D:21 for the Exploration Theme Session D: Comparative Marine Ecosystem of the Sea (ICES) Structure and Function: Descriptors and Characteristics Fronts in the World Ocean’s Large Marine Ecosystems Igor M. Belkin and Peter C. Cornillon Abstract. Oceanic fronts shape marine ecosystems; therefore front mapping and characterization is one of the most important aspects of physical oceanography. Here we report on the first effort to map and describe all major fronts in the World Ocean’s Large Marine Ecosystems (LMEs). Apart from a geographical review, these fronts are classified according to their origin and physical mechanisms that maintain them. This first-ever zero-order pattern of the LME fronts is based on a unique global frontal data base assembled at the University of Rhode Island. Thermal fronts were automatically derived from 12 years (1985-1996) of twice-daily satellite 9-km resolution global AVHRR SST fields with the Cayula-Cornillon front detection algorithm. These frontal maps serve as guidance in using hydrographic data to explore subsurface thermohaline fronts, whose surface thermal signatures have been mapped from space. Our most recent study of chlorophyll fronts in the Northwest Atlantic from high-resolution 1-km data (Belkin and O’Reilly, 2007) revealed a close spatial association between chlorophyll fronts and SST fronts, suggesting causative links between these two types of fronts. Keywords: Fronts; Large Marine Ecosystems; World Ocean; sea surface temperature. Igor M. Belkin: Graduate School of Oceanography, University of Rhode Island, 215 South Ferry Road, Narragansett, Rhode Island 02882, USA [tel.: +1 401 874 6533, fax: +1 874 6728, email: [email protected]]. -

THEME SESSION on North Atlantic Processes (L)
THEME SESSION on North Atlantic Processes (L) ICES CM 2000/L:01 The relation between long-term variations of water temperature in the North Atlantic and Nordic Seas Yu. Bochkov, E. Sentyabov, and A. Karsakov The paper presents the results of estimation of the character and value of the relation between long-term variations of the water thermal state in the North Atlantic and the adjacent Nordic Seas. A close relation is found between large-scale variations of the water thermal state over the study area from the Labrador Sea to the Barents Sea. These long-range relations are of both synchronous and asynchronous character, which permits us to use them with the purpose of forecast. Using data on the sea surface temperature in the North Atlantic (1982–2000), as well as data on the water temperature at the depth of 0–200 m in the Kola Section (the Barents Sea) as the base, a close synchronous relation between interannual variations of the temperature in the Barents Sea and the Gulf Stream areas (positive relation) and the Labrador Current (negative relation) is found. An important factor of formation of the large-scale and long-term variations in climatic systems of the North Atlantic and adjacent seas is the North Atlantic Oscillation. The other character of the relation is revealed when comparing interannual (1959–1999) variations of temperature of the Atlantic waters in the Faeroe-Shetland Channel (Northeast Atlantic) and in the Kola Section (The Barents Sea). Here a close asynchronous relation is found. Temperature variations in the Kola Section are 10–12 months behind those in the Faeroe-Shetland Channel. -
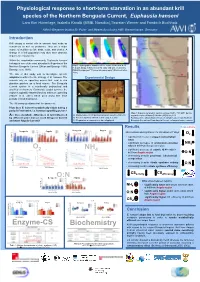
Physiological Response to Short-Term Starvation in an Abundant Krill
Physiological response to short-term starvation in an abundant krill species of the Northern Benguela Current, Euphausia hanseni Lara Kim Hünerlage, Isabella Kandjii (MME, Namibia),Thorsten Werner and Friedrich Buchholz Alfred-Wegener-Institut für Polar- und Meeresforschung AWI, Bremerhaven, Germany Introduction Krill occupy a central role in oceanic food webs as consumers as well as producers. They are a major source of nutrition to fish, birds, seals, and whales. A change in a krill population may thus have dramatic impacts on ecosystems. Within the zooplankton community, Euphausia hanseni belongs to one of the most abundant krill species of the Northern Benguela Current (Olivar and Barange 1990; Map 1: Hydrographic situation off the coast of Namibia at 20 m depth. Image is based on CTD data and was created by Barange et al. 1991). GENUS-subproject “Physical Oceanography” (Mohrholz et al. 2011). The aim of this study was to investigate specific adaptations within the life strategy of E. hanseni. The Experimental Design animals rely on upwelling pulses that lead to rich plankton patches as a food source. The Benguela Current system is a nutritionally poly-pulsed and stratified environment. During late austral summer, the region is typically characterized by minimum upwelling A (Hagen et al. 2001) which goes along with short periods of food deprivation. The following questions shall be answered: How does E. hanseni metabolically adjust during a period of starvation, i.e. between upwelling pulses? C B Map 2: Stations sampled in austral summer 30.01- 7.08.2011 during Are there metabolic differences in krill influenced A) Maintenance of krill during starvation experiment (n=48) research cruise of Maria S. -

St Helena Bay
The variability of retention in St Helena Bay Anathi Manyakanyaka MNYANA002 Supervisors: Dr Jenifer Jackson-Veitch (SAEON) and A/Prof Mathieu Rouault (UCT) A minor dissertation submitted in partial fulfilment of the requirements for the degree of Master of Science in Applied Ocean Sciences of the University of Cape Town University of Cape Town Department of Oceanography Faculty of Science Submitted February 2020 The copyright of this thesis vests in the author. No quotation from it or information derived from it is to be published without full acknowledgement of the source. The thesis is to be used for private study or non- commercial research purposes only. Published by the University of Cape Town (UCT) in terms of the non-exclusive license granted to UCT by the author. University of Cape Town Declaration I know that plagiarism is wrong. Plagiarism is to use another’s work and pretend that it is one’s own. I have used the Harvard Style referencing convention for citation and referencing. Each contribution to, and quotation in, this thesis from the work(s) of other people has been attributed and has been cited and referenced. This thesis is my own work. I have not allowed, and will not allow, anyone to copy my work with the intention of passing it off as his or her own work. Signature ______________________________ Date 06 February 2020 _________________ 1 | Page Abstract The circulation in St Helena Bay and the variability of the retention of the Bay are investigated using seasonal climatologies of the Regional Ocean Modelling System (ROMS). While retention has been studied biologically, the seasonality of the hydrodynamics contributing to the retention have received less attention. -

Characteristics of Intermediate Water Flow in the Benguela Current As
Deep-Sea Research II 50 (2003) 87–118 Characteristics of intermediate water flow in the Benguela current as measured with RAFOS floats P.L. Richardsona,*, S.L. Garzolib a Department of Physical Oceanography, Woods Hole Oceanographic Institution, 360 Woods Hole Road, Woods Hole, MA 02543, 3 Water Street, P.O. Box 721, USA b Atlantic Oceanographic and Meteorological Laboratory, NOAA, 4301 Rickenbacker Causeway, Miami, FL 33149, USA Received 28 September 2001; accepted 26 July 2002 Abstract Seven floats (not launched in rings) crossed over the mid-Atlantic Ridge in the Benguela extension with a mean westward velocity of around 2 cm=s between 22S and 35S. Two Agulhas rings crossed over the mid-Atlantic Ridge with a mean velocity of 5:7cm=s toward 2851: This implies they translated at around 3:8cm=s through the background velocity field near 750 m: The boundaries of the Benguela Current extension were clearly defined from the observations. At 750 m the Benguela extension was bounded on the south by 35S and the north by an eastward current located between 18S and 21S. Other recent float measurements suggest that this eastward current originates near the Trindade Ridge close to the western boundary and extends across most of the South Atlantic, limiting the Benguela extension from flowing north of around 20S. The westward transport of the Benguela extension was estimated to be 15 Sv by integrating the mean westward velocities from 22S to 35S and multiplying by the 500 m estimated thickness of intermediate water. Roughly 1.5 Sv of this are transported by the B3 Agulhas rings that cross the mid-Atlantic Ridge each year (as observed with altimetry). -

Global Ocean Meridional Overturning
2550 JOURNAL OF PHYSICAL OCEANOGRAPHY VOLUME 37 NOTES AND CORRESPONDENCE Global Ocean Meridional Overturning RICK LUMPKIN Physical Oceanography Division, NOAA/Atlantic Oceanographic and Meteorological Laboratory, Miami, Florida KEVIN SPEER Department of Oceanography, The Florida State University, Tallahassee, Florida (Manuscript received 9 August 2005, in final form 9 January 2007) ABSTRACT A decade-mean global ocean circulation is estimated using inverse techniques, incorporating air–sea fluxes of heat and freshwater, recent hydrographic sections, and direct current measurements. This infor- mation is used to determine mass, heat, freshwater, and other chemical transports, and to constrain bound- ary currents and dense overflows. The 18 boxes defined by these sections are divided into 45 isopycnal (neutral density) layers. Diapycnal transfers within the boxes are allowed, representing advective fluxes and mixing processes. Air–sea fluxes at the surface produce transfers between outcropping layers. The model obtains a global overturning circulation consistent with the various observations, revealing two global-scale meridional circulation cells: an upper cell, with sinking in the Arctic and subarctic regions and upwelling in the Southern Ocean, and a lower cell, with sinking around the Antarctic continent and abyssal upwelling mainly below the crests of the major bathymetric ridges. 1. Introduction (WGASF 2001; Garnier et al. 2000; Josey 2001; Josey et al. 1999). The global pattern of wind and heat gain and Wind, and heat and freshwater fluxes at the ocean loss in these products is qualitatively consistent in the surface are, together with tidal and other energy Northern Hemisphere where the ocean gains heat in sources, responsible for the global ocean circulation, the Tropics and loses large amounts of heat in the mixing, and the formation of a broad range of water northern North Atlantic. -

Lecture 4: OCEANS (Outline)
LectureLecture 44 :: OCEANSOCEANS (Outline)(Outline) Basic Structures and Dynamics Ekman transport Geostrophic currents Surface Ocean Circulation Subtropicl gyre Boundary current Deep Ocean Circulation Thermohaline conveyor belt ESS200A Prof. Jin -Yi Yu BasicBasic OceanOcean StructuresStructures Warm up by sunlight! Upper Ocean (~100 m) Shallow, warm upper layer where light is abundant and where most marine life can be found. Deep Ocean Cold, dark, deep ocean where plenty supplies of nutrients and carbon exist. ESS200A No sunlight! Prof. Jin -Yi Yu BasicBasic OceanOcean CurrentCurrent SystemsSystems Upper Ocean surface circulation Deep Ocean deep ocean circulation ESS200A (from “Is The Temperature Rising?”) Prof. Jin -Yi Yu TheThe StateState ofof OceansOceans Temperature warm on the upper ocean, cold in the deeper ocean. Salinity variations determined by evaporation, precipitation, sea-ice formation and melt, and river runoff. Density small in the upper ocean, large in the deeper ocean. ESS200A Prof. Jin -Yi Yu PotentialPotential TemperatureTemperature Potential temperature is very close to temperature in the ocean. The average temperature of the world ocean is about 3.6°C. ESS200A (from Global Physical Climatology ) Prof. Jin -Yi Yu SalinitySalinity E < P Sea-ice formation and melting E > P Salinity is the mass of dissolved salts in a kilogram of seawater. Unit: ‰ (part per thousand; per mil). The average salinity of the world ocean is 34.7‰. Four major factors that affect salinity: evaporation, precipitation, inflow of river water, and sea-ice formation and melting. (from Global Physical Climatology ) ESS200A Prof. Jin -Yi Yu Low density due to absorption of solar energy near the surface. DensityDensity Seawater is almost incompressible, so the density of seawater is always very close to 1000 kg/m 3. -

Atlantic Ocean Equatorial Currents
188 ATLANTIC OCEAN EQUATORIAL CURRENTS ATLANTIC OCEAN EQUATORIAL CURRENTS S. G. Philander, Princeton University, Princeton, Centered on the equator, and below the westward NJ, USA surface Sow, is an intense eastward jet known as the Equatorial Undercurrent which amounts to a Copyright ^ 2001 Academic Press narrow ribbon that precisely marks the location of doi:10.1006/rwos.2001.0361 the equator. The undercurrent attains speeds on the order of 1 m s\1 has a half-width of approximately Introduction 100 km; its core, in the thermocline, is at a depth of approximately 100 m in the west, and shoals to- The circulations of the tropical Atlantic and PaciRc wards the east. The current exists because the west- Oceans have much in common because similar trade ward trade winds, in addition to driving divergent winds, with similar seasonal Suctuations, prevail westward surface Sow (upwelling is most intense at over both oceans. The salient features of these circu- the equator), also maintain an eastward pressure lations are alternating bands of eastward- and west- force by piling up the warm surface waters in the ward-Sowing currents in the surface layers (see western side of the ocean basin. That pressure force Figure 1). Fluctuations of the currents in the two is associated with equatorward Sow in the thermo- oceans have similarities not only on seasonal but cline because of the Coriolis force. At the equator, even on interannual timescales; the Atlantic has where the Coriolis force vanishes, the pressure force a phenomenon that is the counterpart of El Ninoin is the source of momentum for the eastward Equa- the PaciRc. -

EASTERN BOUNDARY CIRCULATION and HYDROGRAPHY OFF ANGOLA Building Angolan Oceanographic Capacities
EASTERN BOUNDARY CIRCULATION AND HYDROGRAPHY OFF ANGOLA Building Angolan Oceanographic Capacities P. TCHIPALANGA, M. DENGLER, P. BRANdt, R. KOPTE, M. MACUÉRIA, P. COELHO, M. OSTROWSKI, AND N. S. KEENLYSIDE The seasonal circulation and interannual hydrographic variability off the coast of Angola is revealed by biannual research cruise data (1995–2017) from the Nansen Programme. ngola is located at the Atlantic coast in south- development. Currently, the fishing sector is third western Africa between 5° and 17°20ʹS, with in importance to the national economy after the oil Aborders to the Democratic Republic of Congo in and mining industries and supplies about 25% of the the north and to Namibia in the south. Its coastline total animal protein intake of the Angolan popula- stretches over a distance of 1,600 km. The Angolan tion (FAO 2011). However, fish resources are found territorial waters support a highly productive ecosys- to be affected by climate variability and changes tem. Seasonal upwelling occurs in large parts of its of the eastern boundary upwelling ecosystems coastal zone during austral winter (Fig. 1). Coupled (e.g., Gammelsrød et al. 1998; Parrish et al. 2000; with a dense coastal population, this marine ecosys- Lehodey et al. 2006; Gruber 2011). There is urgent tem plays a key socioeconomic role in the country’s need to understand these impacts to help sustainable AFFILIATIONS: TCHIPALANGA—Departamento do Ambiente Norway; KEENLYSIDE—Geophysical Institute, Bjerknes Centre for e Ecossistemas Aquáticos, Instituto Nacional de Investigação Climate Research, University of Bergen, Bergen, Norway Pesqueira, Moçâmedes, Angola; DENGLER AND KOPTE—Physical Ocean- CORRESPONDING AUTHOR: Marcus Dengler, ography, Ocean Circulation and Climate Dynamics, GEOMAR [email protected] Helmholtz Centre for Ocean Research, Kiel, Germany; BRANdt— The abstract for this article can be found in this issue, following the table Physical Oceanography, Ocean Circulation and Climate Dynamics, of contents. -

Upper-Level Circulation in the South Atlantic Ocean
Prog. Oceanog. Vol. 26, pp. 1-73, 1991. 0079 - 6611/91 $0.00 + .50 Printed in Great Britain. All fights reserved. © 1991 Pergamon Press pie Upper-level circulation in the South Atlantic Ocean RAY G. P~-rwtSON and LOTHAR Sa~AMMA lnstitut fiir Meereskunde an der Universitiit Kiel, Diisternbrooker Weg 20, 2300 Kiel 1, F.R.G. Abstract - In this paper we present a literature survey of the South Atlantic's climate and its oceanic upper-layer circulation and meridional beat transport. The opening section deals with climate and is focused upon those elements having greatest oceanic relevance, i.e., distributions of atmospheric sea level pressure, the wind fields they produce, and the net surface energy fluxes. The various geostrophic currents comprising the upper-level general circulation are then reviewed in a manner organized around the subtropical gyre, beginning off southern Africa with the Agulhas Current Retroflection and then progressing to the Benguela Current, the equatorial current system and circulation in the Angola Basin, the large-scale variability and interannual warmings at low latitudes, the Brazil Current, the South Atlantic Cmrent, and finally to the Antarctic Circumpolar Current system in which the Falkland (Malvinas) Current is included. A summary of estimates of the meridional heat transport at various latitudes in the South Atlantic ends the survey. CONTENTS 1. Introduction 2 2. Climatic Elements 2 3. Subtropical and Equatorial Circulation 11 3.1. Agulhas Current Retroflection 11 3.2. Benguela Cmrent 16 3.3. Equatorial Cttrrents 18 3.3.1. Components of the system 18 3.3.2. Angola Basin circulation 26 3.3.3. -

Benguela Current Large Marine Ecosystem (LME)
OPTIONAL ANNEXES Identifiers: Project Number: PIMS: 0096 UNDP: RAF00G31 Project Name: Implementation of the Strategic Action Program (SAP) Toward Achievement of the Integrated Management of the Benguela Current Large Marine Ecosystem (LME) Annex 5: Transboundary Diagnostic Analysis Annex 6: Strategic Action Programme Annex 7: Summary of the Functions and Responsibilities of the Interim Benguela Current Commission (IBCC) Annex 8: Thematic Reports Prepared During the PDF-B Project Phase Annex 9: Stakeholders Involvement Description and List of the Stakeholders Participants 2 ANNEX H BENGUELA CURRENT LARGE MARINE ECOSYSTEM PROGRAMME (BCLME) A regional commitment to the sustainable integrated management of the Benguela Current Large Marine Ecosystem by Angola, Namibia and South Africa TRANSBOUNDARY DIAGNOSTIC ANALYSIS (TDA) UNDP WINDHOEK, OCTOBER 1999 3 TABLE OF CONTENTS Page Background and Introduction 1 * The Benguela: a unique environment 1 * Fragmented management: a legacy of the 2 colonial and political past * The need for international action 3 * The success story of BENEFIT 4 * The emerging BCLME Programme 6 * What has been achieved 6 * Towards a sustainable future: the next steps 8 Users Guide to the Transboundary Diagnostic Analysis 11 * Definitions and TDA objective 11 * Design of the TDA 11 (a) Level One: Synthesis (b) Level Two: Specifics * More information 12 BCLME Transboundary Diagnostic Analysis 13 * Geographic scope and ecosystem boundaries 13 * Level One: Synthesis 14 * Synthesis Matrix 16 * Level Two: Overview 17 * Analysis