Chapter 12 Breeding Bird Communities
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Birds of the East Texas Baptist University Campus with Birds Observed Off-Campus During BIOL3400 Field Course
Birds of the East Texas Baptist University Campus with birds observed off-campus during BIOL3400 Field course Photo Credit: Talton Cooper Species Descriptions and Photos by students of BIOL3400 Edited by Troy A. Ladine Photo Credit: Kenneth Anding Links to Tables, Figures, and Species accounts for birds observed during May-term course or winter bird counts. Figure 1. Location of Environmental Studies Area Table. 1. Number of species and number of days observing birds during the field course from 2005 to 2016 and annual statistics. Table 2. Compilation of species observed during May 2005 - 2016 on campus and off-campus. Table 3. Number of days, by year, species have been observed on the campus of ETBU. Table 4. Number of days, by year, species have been observed during the off-campus trips. Table 5. Number of days, by year, species have been observed during a winter count of birds on the Environmental Studies Area of ETBU. Table 6. Species observed from 1 September to 1 October 2009 on the Environmental Studies Area of ETBU. Alphabetical Listing of Birds with authors of accounts and photographers . A Acadian Flycatcher B Anhinga B Belted Kingfisher Alder Flycatcher Bald Eagle Travis W. Sammons American Bittern Shane Kelehan Bewick's Wren Lynlea Hansen Rusty Collier Black Phoebe American Coot Leslie Fletcher Black-throated Blue Warbler Jordan Bartlett Jovana Nieto Jacob Stone American Crow Baltimore Oriole Black Vulture Zane Gruznina Pete Fitzsimmons Jeremy Alexander Darius Roberts George Plumlee Blair Brown Rachel Hastie Janae Wineland Brent Lewis American Goldfinch Barn Swallow Keely Schlabs Kathleen Santanello Katy Gifford Black-and-white Warbler Matthew Armendarez Jordan Brewer Sheridan A. -

The Status and Occurrence of Black Phoebe (Sayornis Nigricans) in British Columbia
The Status and Occurrence of Black Phoebe (Sayornis nigricans) in British Columbia. By Rick Toochin. Introduction and Distribution The Black Phoebe (Sayornis nigricans) is a small passerine belonging to the tyrant-flycatcher family. The Black Phoebe occurs as a year-round resident throughout most of its range; however, its northern populations are partially migratory (Wahl et al. 2005). It is a species found throughout the year from southwestern Oregon south, through California including the Baja Peninsula (excluding the central regions of the Peninsula), east through Arizona, New Mexico, southern Colorado, west Texas, south through Mexico, Central America to Panama (excluding El Salvador) and in South America from the coastal mountains of Venezuela, through Colombia, Ecuador, and Peru, to western Bolivia and northwestern Argentina (Sibley 2000, Howell and Webb 2010, Hoyo et al. 2006). In the past couple of decades the Black Phoebe has been slowly expanding its known range northward into northern Oregon and southern Washington where it is still considered a very rare visitor, but with records increasing every year (Wahl et al. 2005, WBRC 2012). The Black Phoebe has been recorded from Idaho, Nevada, Utah, southern Oklahoma and Florida (Sibley 2000). The Black Phoebe is an accidental visitor to south-central Alaska (Gibson et al. 2013). In British Columbia this species is considered a casual visitor but Provincial records, like those of Washington State, are on the rise and the status of this species in British Columbia could change in the near future. Identification and Similar Species The Black Phoebe has a huge range that encompasses two continents. -

Ecology, Morphology, and Behavior in the New World Wood Warblers
Ecology, Morphology, and Behavior in the New World Wood Warblers A dissertation presented to the faculty of the College of Arts and Sciences of Ohio University In partial fulfillment of the requirements for the degree Doctor of Philosophy Brandan L. Gray August 2019 © 2019 Brandan L. Gray. All Rights Reserved. 2 This dissertation titled Ecology, Morphology, and Behavior in the New World Wood Warblers by BRANDAN L. GRAY has been approved for the Department of Biological Sciences and the College of Arts and Sciences by Donald B. Miles Professor of Biological Sciences Florenz Plassmann Dean, College of Arts and Sciences 3 ABSTRACT GRAY, BRANDAN L., Ph.D., August 2019, Biological Sciences Ecology, Morphology, and Behavior in the New World Wood Warblers Director of Dissertation: Donald B. Miles In a rapidly changing world, species are faced with habitat alteration, changing climate and weather patterns, changing community interactions, novel resources, novel dangers, and a host of other natural and anthropogenic challenges. Conservationists endeavor to understand how changing ecology will impact local populations and local communities so efforts and funds can be allocated to those taxa/ecosystems exhibiting the greatest need. Ecological morphological and functional morphological research form the foundation of our understanding of selection-driven morphological evolution. Studies which identify and describe ecomorphological or functional morphological relationships will improve our fundamental understanding of how taxa respond to ecological selective pressures and will improve our ability to identify and conserve those aspects of nature unable to cope with rapid change. The New World wood warblers (family Parulidae) exhibit extensive taxonomic, behavioral, ecological, and morphological variation. -

Bird List of San Bernardino Ranch in Agua Prieta, Sonora, Mexico
Bird List of San Bernardino Ranch in Agua Prieta, Sonora, Mexico Melinda Cárdenas-García and Mónica C. Olguín-Villa Universidad de Sonora, Hermosillo, Sonora, Mexico Abstract—Interest and investigation of birds has been increasing over the last decades due to the loss of their habitats, and declination and fragmentation of their populations. San Bernardino Ranch is located in the desert grassland region of northeastern Sonora, México. Over the last decade, restoration efforts have tried to address the effects of long deteriorating economic activities, like agriculture and livestock, that used to take place there. The generation of annual lists of the wildlife (flora and fauna) will be important information as we monitor the progress of restoration of this area. As part of our professional training, during the summer and winter (2011-2012) a taxonomic list of bird species of the ranch was made. During this season, a total of 85 species and 65 genera, distributed over 30 families were found. We found that five species are on a risk category in NOM-059-ECOL-2010 and 76 species are included in the Red List of the International Union for Conservation of Nature (IUCN). It will be important to continue this type of study in places that are at- tempting restoration and conservation techniques. We have observed a huge change, because of restoration activities, in the lands in the San Bernardino Ranch. Introduction migratory (Villaseñor-Gómez et al., 2010). Twenty-eight of those species are considered at risk on a global scale, and are included in Birds represent one of the most remarkable elements of our en- the Red List of the International Union for Conservation of Nature vironment, because they’re easy to observe and it’s possible to find (IUCN). -

Phylloscopus Sibilatrix
Phylloscopus sibilatrix -- (Bechstein, 1793) ANIMALIA -- CHORDATA -- AVES -- PASSERIFORMES -- SYLVIIDAE Common names: Wood Warbler; Pouillot siffleur European Red List Assessment European Red List Status LC -- Least Concern, (IUCN version 3.1) Assessment Information Year published: 2015 Date assessed: 2015-03-31 Assessor(s): BirdLife International Reviewer(s): Symes, A. Compiler(s): Ashpole, J., Burfield, I., Ieronymidou, C., Pople, R., Wheatley, H. & Wright, L. Assessment Rationale European regional assessment: Least Concern (LC) EU27 regional assessment: Least Concern (LC) At both European and EU27 scales this species has an extremely large range, and hence does not approach the thresholds for Vulnerable under the range size criterion (Extent of Occurrence 10% in ten years or three generations, or with a specified population structure). The population trend appears to be stable, and hence the species does not approach the thresholds for Vulnerable under the population trend criterion (30% decline over ten years or three generations). For these reasons the species is evaluated as Least Concern within both Europe and the EU27. Occurrence Countries/Territories of Occurrence Native: Albania; Andorra; Armenia; Austria; Azerbaijan; Belarus; Belgium; Bosnia and Herzegovina; Bulgaria; Croatia; Cyprus; Czech Republic; Denmark; Estonia; Finland; France; Georgia; Germany; Greece; Hungary; Ireland, Rep. of; Italy; Latvia; Liechtenstein; Lithuania; Luxembourg; Macedonia, the former Yugoslav Republic of; Malta; Moldova; Montenegro; Netherlands; Norway; Poland; Romania; Russian Federation; Serbia; Slovakia; Slovenia; Spain; Canary Is. (to ES); Sweden; Switzerland; Turkey; Ukraine; United Kingdom Vagrant: Faroe Islands (to DK); Iceland; Portugal; Gibraltar (to UK) Population The European population is estimated at 7,060,000-11,100,000 pairs, which equates to 14,100,000-22,100,000 mature individuals. -

An Apparent Hybrid Black × Eastern Phoebe from Colorado Nathan D
AN APPARENT HYBRID BLACK × EASTERN PHOEBE FROM COLORADO NATHAN D. PIEPLOW, University of Colorado, 317 UCB, Boulder, Colorado 80309; [email protected] TONY LEUKERING, P. O. Box 660, Brighton, Colorado 80601 ELAINE COLEY, 290 Riker Court, Loveland, Colorado 80537 ABSTRACT: From 21 April to 11 May 2007 an apparent hybrid male Black × Eastern Phoebe (Sayornis nigricans × S. phoebe) was observed in Loveland, Larimer County, Colorado. The bird’s plumage was intermediate between the species, with paler upperparts, darker flanks, and a less distinct border between dark and white on the breast than expected on the Black Phoebe and darker upperparts, less head/back contrast, and a darker and more sharply demarcated upper breast than expected on the Eastern. Sonograms of the bird’s territorial song show numerous characteristics intermediate between the typical songs of the two species. Although the Colorado bird provides the first strong evidence of hybridization in Sayornis, other sightings suggest the Black and Eastern Phoebes may have hybridized on other recent occa- sions. Range expansions of both species may increase the frequency of hybridization in the future. The Black (Sayornis nigricans) and Eastern (S. phoebe) Phoebes are similar medium-sized flycatchers that historically occupied allopatric breed- ing ranges. The Eastern Phoebe is a common breeding species of mixed woodlands, riparian gallery forest, and human-altered habitats, nesting on cliffs, banks, and man-made structures in the eastern two-thirds of the lower 48 states and in Canada from Nova Scotia west and north to southwestern Nunavut and northeastern British Columbia (Weeks 1994). The Great Plains were probably a barrier to the species’ range expansion until channelization and stabilization of river banks encouraged the growth of extensive gallery forests along east-flowing rivers, particularly the South Platte andA rkansas (Knopf 1991). -
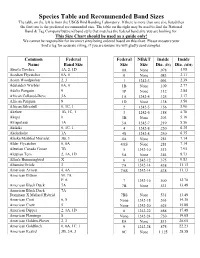
Species Table and Recommended Band Sizes the Table on the Left Is from the USGS Bird Banding Laboratory
Species Table and Recommended Band Sizes The table on the left is from the USGS Bird Banding Laboratory. If there is more than one size listed then the first one is the preferred recommended size. The table on the right may be used to find the National Band & Tag Company butt-end band style that matches the federal band size you are looking for. This Size Chart should be used as a guide only! We cannot be responsible for incorrect sizes being ordered based on this chart. Please measure your bird’s leg for accurate sizing, if you are unsure we will gladly send samples. Common Federal Federal NB&T Inside Inside Name Band Size Size Size Dia. (IN) Dia. (MM) Abert's Towhee 1A, 2, 1D 0A None .078 1.98 Acadian Flycatcher 0A, 0 0 None .083 2.11 Acorn Woodpecker 2, 3 1 1242-3 .094 2.39 Adelaide's Warbler 0A, 0 1B None .109 2.77 Adelie Penguin 9 1P None .112 2.84 African Collared-Dove 3A 1A 1242-4 .125 3.17 African Penguin 9 1D None .138 3.50 African Silverbill 0, 1C, 1 2 1242-5 .156 3.96 Akekee 1B, 1C, 1 3 1242-6 .188 4.78 Akepa 0 3B None .203 5.16 Akiapolaau 1A 3A 1242-7 .219 5.56 Akikiki 0, 1C, 1 4 1242-8 .250 6.35 Akohekohe 1A 4S 1242-8 .250 6.35 Alaska Marbled Murrelet 3B, 3 4A None .281 7.14 Alder Flycatcher 0, 0A 4AS None .281 7.14 Aleutian Canada Goose 7B 5 1242-10 .313 7.95 Aleutian Tern 2, 1A, 1D 5A None .344 8.73 Allen's Hummingbird X 6 1242-12 .375 9.53 Altamira Oriole 3 7A 1242-14 .438 11.13 American Avocet 4, 4A 7AS 1242-14 .438 11.13 American Bittern M: 7A F: 6 7 1242-16 .500 12.70 American Black Duck 7A 7B None .531 13.49 American -

Learn About Texas Birds Activity Book
Learn about . A Learning and Activity Book Color your own guide to the birds that wing their way across the plains, hills, forests, deserts and mountains of Texas. Text Mark W. Lockwood Conservation Biologist, Natural Resource Program Editorial Direction Georg Zappler Art Director Elena T. Ivy Educational Consultants Juliann Pool Beverly Morrell © 1997 Texas Parks and Wildlife 4200 Smith School Road Austin, Texas 78744 PWD BK P4000-038 10/97 All rights reserved. No part of this work covered by the copyright hereon may be reproduced or used in any form or by any means – graphic, electronic, or mechanical, including photocopying, recording, taping, or information storage and retrieval systems – without written permission of the publisher. Another "Learn about Texas" publication from TEXAS PARKS AND WILDLIFE PRESS ISBN- 1-885696-17-5 Key to the Cover 4 8 1 2 5 9 3 6 7 14 16 10 13 20 19 15 11 12 17 18 19 21 24 23 20 22 26 28 31 25 29 27 30 ©TPWPress 1997 1 Great Kiskadee 16 Blue Jay 2 Carolina Wren 17 Pyrrhuloxia 3 Carolina Chickadee 18 Pyrrhuloxia 4 Altamira Oriole 19 Northern Cardinal 5 Black-capped Vireo 20 Ovenbird 6 Black-capped Vireo 21 Brown Thrasher 7Tufted Titmouse 22 Belted Kingfisher 8 Painted Bunting 23 Belted Kingfisher 9 Indigo Bunting 24 Scissor-tailed Flycatcher 10 Green Jay 25 Wood Thrush 11 Green Kingfisher 26 Ruddy Turnstone 12 Green Kingfisher 27 Long-billed Thrasher 13 Vermillion Flycatcher 28 Killdeer 14 Vermillion Flycatcher 29 Olive Sparrow 15 Blue Jay 30 Olive Sparrow 31 Great Horned Owl =female =male Texas Birds More kinds of birds have been found in Texas than any other state in the United States: just over 600 species. -

Eastern Wood-Pewee,Contopus Virens
COSEWIC Assessment and Status Report on the Eastern Wood-pewee Contopus virens in Canada Photo: courtesy Carl Savignac SPECIAL CONCERN 2012 COSEWIC status reports are working documents used in assigning the status of wildlife species suspected of being at risk. This report may be cited as follows: COSEWIC. 2012. COSEWIC assessment and status report on the Eastern Wood-pewee Contopus virens in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. x + 39 pp. (www.registrelep-sararegistry.gc.ca/default_e.cfm). Production note: COSEWIC would like to acknowledge Carl Savignac for writing the status report on the Eastern Wood- pewee, Contopus virens, in Canada, prepared under contract with Environment Canada. The report was overseen and edited by Jon McCracken, COSEWIC Birds Specialist Subcommittee Co-chair. For additional copies contact: COSEWIC Secretariat c/o Canadian Wildlife Service Environment Canada Ottawa, ON K1A 0H3 Tel.: 819-953-3215 Fax: 819-994-3684 E-mail: COSEWIC/[email protected] http://www.cosewic.gc.ca Également disponible en français sous le titre Ếvaluation et Rapport de situation du COSEPAC sur le Pioui de l’Est (Contopus virens) au Canada. Cover illustration/photo: Eastern Wood-pewee — Photo: courtesy Carl Savignac. Her Majesty the Queen in Right of Canada, 2013. Catalogue No. CW69-14/663-2013E-PDF ISBN 978-1-100-22136-6 Recycled paper COSEWIC Assessment Summary Assessment Summary – November 2012 Common name Eastern Wood-pewee Scientific name Contopus virens Status Special Concern Reason for designation This species is one of the most common and widespread songbirds associated with North America’s eastern forests. -

A Bird's Eye View of the Forest
A BIRD’S EYE VIEW OF THE FOREST: HOW DOES CANOPY OPENNESS AFFECT CANOPY SONGBIRDS? A Thesis Presented in Partial Fulfillment of the Requirements for the Degree Master of Science in the Graduate School of The Ohio State University By Felicity L. Newell Graduate Program in Natural Resources ***** The Ohio State University 2010 Master’s Committee: Dr. Amanda Rodewald, Advisor Dr. Roger Williams Dr. Paul Rodewald Copyright Felicity L. Newell 2010 ABSTRACT Today, the majority of eastern deciduous forests in North America are closed- canopy mature second-growth. In contrast, an open forest canopy is thought to have been important in presettlement forest ecosystems. Changes in forest structure are of special concern given that oaks ( Quercus spp.) cannot regenerate effectively in closed-canopy stands. Although oaks have historically dominated many eastern forests, forest composition is changing due to anthropogenic impacts on disturbance regimes. Consequently, many forest management efforts now aim to promote oak regeneration and restore forest structure, which ultimately will require a more complete understanding of the ecological processes associated with canopy disturbance. Partial harvesting (e.g. shelterwoods) provides one way to examine the importance of canopy openness. My research examined the link between canopy openness and birds that nest and forage in the forest canopy. Specifically I examined the extent to which (1) canopy songbirds select particular canopy features and (2) canopy structure affects reproductive success. I compared measures of preference (abundance, settlement patterns, site fidelity, and age distributions) and fitness (e.g. nesting success) between recent shelterwood harvests and unharvested second-growth stands in mixed-oak forests of southern Ohio. -

Killing Behaviour of Adult Brood Parasites
1 SPECIAL: ANECDOTES IN ANIMAL BEHAVIOUR 2 Killing behaviour of adult brood parasites 3 Šulc M.1,*, Štětková G.1,2, Jelínek V. 1, Czyż B.3, Dyrcz A.3, Karpińska O.4, 4 Kamionka-Kanclerska K.4, Rowiński P.4, Maziarz M.5, Gruszczyński A.5, Hughes A.E.6, 5 Honza M. 1 61Institute of Vertebrate Biology of the Czech Academy of Sciences, Brno, Czech Republic 72Department of Botany and Zoology, Faculty of Sciences, Masaryk University, Brno, Czech 8Republic 93Department of Behavioural Ecology, University of Wrocław, Wrocław, Poland 104Department of Forest Zoology and Wildlife Management, Warsaw University of Life 11Sciences (SGGW), Warsaw, Poland 125Museum and Institute of Zoology, Polish Academy of Sciences, Warsaw, Poland 136Department of Psychology, University of Essex, Colchester, U.K. 14*corresponding author: Michal Šulc ([email protected]) 15 16Summary 17Decades of studies have revealed the striking adaptations of avian brood parasites for 18their unique reproductive lifestyle. Several have reported that adult brood parasites 19sometimes kill host nestlings, although the reasons for this behaviour remain unclear. 20Using continuous video-recording and camera traps, we observed the same behaviour in 21the common cuckoo Cuculus canorus, showing that both host and parasite nestlings can 22be killed. The latter has never previously been observed in any avian brood parasite. 23Here, we review this phenomenon and discuss possible explanations. 24 25Keywords: nestling infanticide, chick ejection, farming, Mafia, co-evolution 26 27Main text 28Infants are an obvious target for extermination because of their vulnerability, therefore they 29are often exploited by various predators (Weidinger 2009). Apart from this relatively simple 30predator–prey relationship, there is also the phenomenon of infanticide when a young 31offspring is killed by an adult animal of the same species. -

Herefore Takes Precedence
Introduction I have endeavored to keep typos, errors etc in this list to a minimum, however when you find more I would be grateful if you could mail the details during 2011 to: [email protected]. Grateful thanks to Dick Coombes for the cover images. Joe Hobbs Index The general order of species follows the International Ornithological Congress’ World Bird List. Version Version 1.9 (1 August 2011). Cover Main image: Arctic Warbler. Cotter’s Garden, Cape Clear Island, Co. Cork, Ireland. 9 October 2009. Richard H. Coombes. Vignette: Arctic Warbler. The Waist, Cape Clear Island, Co. Cork, Ireland. 10 October 2009. Richard H. Coombes. Species Page No. Alpine Leaf Warbler [Phylloscopus occisinensis] 17 Arctic Warbler [Phylloscopus borealis] 24 Ashy-throated Warbler [Phylloscopus maculipennis] 20 Black-capped Woodland Warbler [Phylloscopus herberti] 5 Blyth’s Leaf Warbler [Phylloscopus reguloides] 31 Brooks’ Leaf Warbler [Phylloscopus subviridis] 22 Brown Woodland Warbler [Phylloscopus umbrovirens] 5 Buff-barred Warbler [Phylloscopus pulcher] 19 Buff-throated Warbler [Phylloscopus subaffinis] 17 Canary Islands Chiffchaff [Phylloscopus canariensis] 12 Chiffchaff [Phylloscopus collybita] 8 Chinese Leaf Warbler [Phylloscopus yunnanensis] 20 Claudia’s Leaf Warbler [Phylloscopus claudiae] 31 Davison’s Leaf Warbler [Phylloscopus davisoni] 32 Dusky Warbler [Phylloscopus fuscatus] 15 Eastern Bonelli's Warbler [Phylloscopus orientalis] 14 Eastern Crowned Warbler [Phylloscopus coronatus] 30 Emei Leaf Warbler [Phylloscopus emeiensis] 32 Gansu Leaf Warbler