Boiga Irregularis), a Costly Introduced Pest on Pacific Sli Ands Gordon H
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Pressure and Duration of Constriction in Boa Constrictor Is Influenced by a Simulated Prey Heartbeat Allison Elizabeth Hall Dickinson College
Dickinson College Dickinson Scholar Student Honors Theses By Year Student Honors Theses 5-23-2010 Pressure and Duration of Constriction in Boa Constrictor is Influenced by a Simulated Prey Heartbeat Allison Elizabeth Hall Dickinson College Follow this and additional works at: http://scholar.dickinson.edu/student_honors Part of the Biology Commons Recommended Citation Hall, Allison Elizabeth, "Pressure and Duration of Constriction in Boa Constrictor is Influenced by a Simulated Prey Heartbeat" (2010). Dickinson College Honors Theses. Paper 86. This Honors Thesis is brought to you for free and open access by Dickinson Scholar. It has been accepted for inclusion by an authorized administrator. For more information, please contact [email protected]. Pressure and Duration of Constriction in Boa constrictor is Influenced by a Simulated Prey Heartbeat By Allison E. Hall With the collaboration of Amanda Hayes and Katelyn McCann Submitted in partial fulfillment of Honors Requirements for the Department of Biology Dr. Scott Boback, Supervisor Dr. Charles Zwemer, Supervisor Dr. David Kushner, Reader May 18, 2010 Abstract Constricting prey is energetically costly for snakes and therefore it would be beneficial to minimize this cost. However, the consequences of arresting a constriction event too soon could be deadly. Thus, the duration of constriction is bounded by competing demands to kill prey and conserve energy. Snakes possess mechanoreceptors within their ventral and dorsal skin that are used for detecting approaching predators and prey. This experiment sought to determine whether Boas (Boa constrictor) can sense a simulated heartbeat in their prey. It was predicted that if snakes possess this ability, those constricting rats with a simulated heart would constrict with greater pressure and increased duration than snakes constricting rats without a simulated heartbeat. -
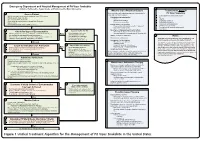
Snake Bite Protocol
Lavonas et al. BMC Emergency Medicine 2011, 11:2 Page 4 of 15 http://www.biomedcentral.com/1471-227X/11/2 and other Rocky Mountain Poison and Drug Center treatment of patients bitten by coral snakes (family Ela- staff. The antivenom manufacturer provided funding pidae), nor by snakes that are not indigenous to the US. support. Sponsor representatives were not present dur- At the time this algorithm was developed, the only ing the webinar or panel discussions. Sponsor represen- antivenom commercially available for the treatment of tatives reviewed the final manuscript before publication pit viper envenomation in the US is Crotalidae Polyva- ® for the sole purpose of identifying proprietary informa- lent Immune Fab (ovine) (CroFab , Protherics, Nash- tion. No modifications of the manuscript were requested ville, TN). All treatment recommendations and dosing by the manufacturer. apply to this antivenom. This algorithm does not con- sider treatment with whole IgG antivenom (Antivenin Results (Crotalidae) Polyvalent, equine origin (Wyeth-Ayerst, Final unified treatment algorithm Marietta, Pennsylvania, USA)), because production of The unified treatment algorithm is shown in Figure 1. that antivenom has been discontinued and all extant The final version was endorsed unanimously. Specific lots have expired. This antivenom also does not consider considerations endorsed by the panelists are as follows: treatment with other antivenom products under devel- opment. Because the panel members are all hospital- Role of the unified treatment algorithm -

Guidelines for Keeping Venomous Snakes in the NT
GUIDELINES FOR KEEPING VENOMOUS SNAKES IN THE NT Venomous snakes are potentially dangerous to humans, and for this reason extreme caution must be exercised when keeping or handling them in captivity. Prospective venomous snake owners should be well informed about the needs and requirements for keeping these animals in captivity. Permits The keeping of protected wildlife in the Northern Territory is regulated by a permit system under the Territory Parks and Wildlife Conservation Act 2006 (TPWC Act). Conditions are included on permits, and the Parks and Wildlife Commission of the Northern Territory (“PWCNT”) may issue infringement notices or cancel permits if conditions are breached. A Permit to Keep Protected Wildlife enables people to legally possess native vertebrate animals in captivity in the Northern Territory. The permit system assists the PWCNT to monitor wildlife kept in captivity and to detect any illegal activities associated with the keeping of, and trade in, native wildlife. Venomous snakes are protected throughout the Northern Territory and may not be removed from the wild without the appropriate licences and permits. People are required to hold a Keep Permit (Category 1–3) to legally keep venomous snakes in the Northern Territory. Premises will be inspected by PWCNT staff to evaluate their suitability prior to any Keep Permit (Category 1– 3) being granted. Approvals may also be required from local councils, the Northern Territory Planning Authority, and the Department of Health and Community Services. Consignment of venomous snakes between the Northern Territory and other States and Territories can only be undertaken with an appropriate import / export permit. There are three categories of venomous snake permitted to be kept in captivity in the Northern Territory: Keep Permit (Category 1) – Mildly Dangerous Venomous Keep Permit (Category 2) – Dangerous Venomous Keep Permit (Category 3) – Highly Dangerous Venomous Venomous snakes must be obtained from a legal source (i.e. -

Reptiles, Birds, and Mammals of Pakin Atoll, Eastern Caroline Islands
Micronesica 29(1): 37-48 , 1996 Reptiles, Birds, and Mammals of Pakin Atoll, Eastern Caroline Islands DONALD W. BUDEN Division Mathematics of and Science, College of Micronesia, P. 0 . Box 159 Kolonia, Polmpei, Federated States of Micronesia 96941. Abstract-Fifteen species of reptiles, 18 birds, and five mammals are recorded from Pakin Atoll. None is endemic to Pakin and all of the residents tend to be widely distributed throughout Micronesia. Intro duced species include four mammals (Rattus exulans, Canis fami/iaris, Fe/is catus, Sus scrofa), the Red Junglefowl (Gallus gal/us) among birds, and at least one lizard (Varanus indicus). Of the 17 indigenous birds, ten are presumed or documented breeding residents, including four land birds, a heron, and five terns. The Micronesian Honeyeater (My=omela rubratra) is the most common land bird, followed closely by the Micro nesian Starling (Aplonis opaca). The vegetation is mainly Cocos forest, considerably modified by periodic cutting of the undergrowth, deliber ately set fires, and the rooting of pigs. Most of the present vertebrate species do not appear to be seriously endangered by present levels of human activity. But the Micronesian Pigeon (Ducula oceanica) is less numerous on the settled islands, probably reflecting increased hunting pressure, and sea turtles (especially Chelonia mydas) and their eggs are harvested indiscriminately . Introduction Terrestrial vertebrates have been poorly studied on many of the remote atolls of Micronesia, and distributional records are lacking or scanty for many islands. The present study documents the occurrence and relative abundance of reptiles, birds, and mammals on Pakin Atoll for the first time. -

BIB 13484.Pdf
Russian Journal of Herpetology Vol. 26, No. 5, 2019, pp. 247 – 260 DOI: 10.30906/1026-2296-2019-26-5-247-260 REAPPRAISAL OF HERPETOFAUNA RECORDED FROM JAFFNA PENINSULA IN NORTHERN SRI LANKA WITH REMARKS ON CONSERVATION, DIVERSITY, AND DISTRIBUTION Majintha Madawala,1 Thilina Surasinghe,2* Anslem De Silva,3 Dinesh Gabadage,4 Madhava Botejue,4 Indika Peabotuwage,5 Dushantha Kandambi,5 and Suranjan Karunarathna5 Submitted January 11, 2017 Jaffna peninsula is quite an unexplored area of Sri Lanka’s lowland dry zone. We constructed a species checklist for all herpetofauna of this area based on a short-term field survey, a comprehensive literature review, museum specimens, and observations made by field herpetologists. Based on 200 × 10 m belt transects, we surveyed herpetofauna both during day and night time, in 10 different types of habitats. The species checklist we compiled comprised 44 species of reptiles (including three nationally threatened, one globally threatened, and eight endemic species) and 15 species of amphibians (including one nationally threatened and three endemic species). Based on published literature, museum specimens, expert opinions, and current field survey, we documented 85 species of herpetofauna in this area. Of this entire list, we were unable to record the presence of 25 species through our field survey. Our field survey documented 18 species that were not previously reported from Jaffna Peninsula. Our study revealed that inland water bodies, cultivated lands, home gardens, and coastal beaches are of high impor- tance for native herpetofauna of Jaffna peninsula. Many human disturbances, such as habitat alterations, vengeful killing, consumption overexploitation, and road mortality are the key threats encountered by herpetofauna in Jaffna. -

Island Biology Island Biology
IIssllaanndd bbiioollooggyy Allan Sørensen Allan Timmermann, Ana Maria Martín González Camilla Hansen Camille Kruch Dorte Jensen Eva Grøndahl, Franziska Petra Popko, Grete Fogtmann Jensen, Gudny Asgeirsdottir, Hubertus Heinicke, Jan Nikkelborg, Janne Thirstrup, Karin T. Clausen, Karina Mikkelsen, Katrine Meisner, Kent Olsen, Kristina Boros, Linn Kathrin Øverland, Lucía de la Guardia, Marie S. Hoelgaard, Melissa Wetter Mikkel Sørensen, Morten Ravn Knudsen, Pedro Finamore, Petr Klimes, Rasmus Højer Jensen, Tenna Boye Tine Biedenweg AARHUS UNIVERSITY 2005/ESSAYS IN EVOLUTIONARY ECOLOGY Teachers: Bodil K. Ehlers, Tanja Ingversen, Dave Parker, MIchael Warrer Larsen, Yoko L. Dupont & Jens M. Olesen 1 C o n t e n t s Atlantic Ocean Islands Faroe Islands Kent Olsen 4 Shetland Islands Janne Thirstrup 10 Svalbard Linn Kathrin Øverland 14 Greenland Eva Grøndahl 18 Azores Tenna Boye 22 St. Helena Pedro Finamore 25 Falkland Islands Kristina Boros 29 Cape Verde Islands Allan Sørensen 32 Tristan da Cunha Rasmus Højer Jensen 36 Mediterranean Islands Corsica Camille Kruch 39 Cyprus Tine Biedenweg 42 Indian Ocean Islands Socotra Mikkel Sørensen 47 Zanzibar Karina Mikkelsen 50 Maldives Allan Timmermann 54 Krakatau Camilla Hansen 57 Bali and Lombok Grete Fogtmann Jensen 61 Pacific Islands New Guinea Lucía de la Guardia 66 2 Solomon Islands Karin T. Clausen 70 New Caledonia Franziska Petra Popko 74 Samoa Morten Ravn Knudsen 77 Tasmania Jan Nikkelborg 81 Fiji Melissa Wetter 84 New Zealand Marie S. Hoelgaard 87 Pitcairn Katrine Meisner 91 Juan Fernandéz Islands Gudny Asgeirsdottir 95 Hawaiian Islands Petr Klimes 97 Galápagos Islands Dorthe Jensen 102 Caribbean Islands Cuba Hubertus Heinicke 107 Dominica Ana Maria Martin Gonzalez 110 Essay localities 3 The Faroe Islands Kent Olsen Introduction The Faroe Islands is a treeless archipelago situated in the heart of the warm North Atlantic Current on the Wyville Thompson Ridge between 61°20’ and 62°24’ N and between 6°15’ and 7°41’ W. -

Occurrence and Distribution of Snake Species in Balochistan Province, Pakistan
Pakistan J. Zool., pp 1-4, 2021. DOI: https://dx.doi.org/10.17582/journal.pjz/20181111091150 Short Communication Occurrence and Distribution of Snake Species in Balochistan Province, Pakistan Saeed Ahmed Essote1, Asim Iqbal1, Muhammad Kamran Taj2*, Asmatullah Kakar1, Imran Taj2, Shahab-ud-Din Kakar1 and Imran Ali 3,4* 1Department of Zoology, University of Balochistan, Quetta 2Center for Advanced Studies in Vaccinology and Biotechnology, University of Balochistan, Quetta, Pakistan. 3Institute of Biochemistry, University of Balochistan, Quetta 4 Plant Biomass Utilization Research Unit, Chulalongkorn University, Bangkok, 10330, Article Information Thailand. Received 11 November 2018 Revised 11 October 2020 Accepted 10 December 2020 ABSTRACT Available online 28 April 2021 Authors’ Contribution The current study was conducted in Zhob, Quetta, Sibi, Kalat, Naseer Abad and Makran Divisions of SAE carried the research with the Balochistan Province. A total of 619 snake specimens representing 6 families, 20 genera and 37 species help of other authors and wrote were collected. The family wise representation among collected specimens has been Boidae (4.6%), the manuscript. AI, MKT and IT Leptotyphlopidae (7.5%), Typhlopidae (10.3%), Elapidae (11.7%), Viperidae (13.4%) and Colubridae helped in the experimental work. AK (52.5%). The percentage of family Boidae, Typhlopidae, Elapidae and Leptotyphlopidae were high in and SDK classified the species and Sibi Division while family Viperidae and Colubridae were dominant in Quetta Division. The family proofread the article. IA helped in arranging contents of the article. Colubridae has been the most dominant in the Province, having ten genera viz., Boiga (6.8%) Coluber (10.1%), Eirenis (2.5%), Lycodon (3.5%), Lytorhynchus (6.1%), Oligodon (4.7%), Natrix (1.7%), (7.5%), Key words Ptyas (2.9 %) Spalerosophis (6.3%) and Psammophis. -

Operational Control of the Brown Tree Snake on Guam
OPERATIONAL CONTROL OF THE BROWN TREE SNAKE ON GUAM THOMAS C. HALL, USDA/APHIS/ADC, 2800 N. Lincoln Blvd., Oklahoma City, Oklahoma 73105. ABSTRACT: An operational control program for brown tree snakes (Boiga i"egularis) on Guam began in April 1993. The program focused on minimizing the dispersal of brown tree snakes to other Pacific islands and the U.S. mainland. During the first year of operation, more than 3,000 snakes were caught within a kilometer of high risk port facilities using traps, detector dogs, and spotlighting. Additionally, habitat modifications and prey-base removal were used to reduce the attractiveness of these facilities to brown tree snakes. Public awareness was also an important part of the program such as the education of cargo packers, shippers, and Customs inspectors who could further minimiu brown tree snake dispersal off-island. Initial control efforts in the program became more efficient with the recognition of brown tree snake characteristics, i.e., it was discovered that perimeter trapping a 5 ha patch of jungle was sufficient to remove most snakes instead of saturating the area with traps. KEY WORDS: brown tree snake, snake control Proc. 17th Vertebr. Pest Conf. (R.M. Timm & A.C. Crabb, Eds.) Published at Univ. of Calif., Davis. 1996. INTRODUCTION methods for resolving the problem. Background Brown tree snakes have caused significant information is given here so that the complexity of this environmental and economic impacts since their problem can be understood. inadvertent introduction to the island of Guam in the late 1940s. Other islands in the Pacific and the U.S. -

Cretaceous Fossil Gecko Hand Reveals a Strikingly Modern Scansorial Morphology: Qualitative and Biometric Analysis of an Amber-Preserved Lizard Hand
Cretaceous Research 84 (2018) 120e133 Contents lists available at ScienceDirect Cretaceous Research journal homepage: www.elsevier.com/locate/CretRes Cretaceous fossil gecko hand reveals a strikingly modern scansorial morphology: Qualitative and biometric analysis of an amber-preserved lizard hand * Gabriela Fontanarrosa a, Juan D. Daza b, Virginia Abdala a, c, a Instituto de Biodiversidad Neotropical, CONICET, Facultad de Ciencias Naturales e Instituto Miguel Lillo, Universidad Nacional de Tucuman, Argentina b Department of Biological Sciences, Sam Houston State University, 1900 Avenue I, Lee Drain Building Suite 300, Huntsville, TX 77341, USA c Catedra de Biología General, Facultad de Ciencias Naturales, Universidad Nacional de Tucuman, Argentina article info abstract Article history: Gekkota (geckos and pygopodids) is a clade thought to have originated in the Early Cretaceous and that Received 16 May 2017 today exhibits one of the most remarkable scansorial capabilities among lizards. Little information is Received in revised form available regarding the origin of scansoriality, which subsequently became widespread and diverse in 15 September 2017 terms of ecomorphology in this clade. An undescribed amber fossil (MCZ Re190835) from mid- Accepted in revised form 2 November 2017 Cretaceous outcrops of the north of Myanmar dated at 99 Ma, previously assigned to stem Gekkota, Available online 14 November 2017 preserves carpal, metacarpal and phalangeal bones, as well as supplementary climbing structures, such as adhesive pads and paraphalangeal elements. This fossil documents the presence of highly specialized Keywords: Squamata paleobiology adaptive structures. Here, we analyze in detail the manus of the putative stem Gekkota. We use Paraphalanges morphological comparisons in the context of extant squamates, to produce a detailed descriptive analysis Hand evolution and a linear discriminant analysis (LDA) based on 32 skeletal variables of the manus. -

Snakes of South-East Asia Including Myanmar, Thailand, Malaysia, Singapore, Sumatra, Borneo, Java and Bali
A Naturalist’s Guide to the SNAKES OF SOUTH-EAST ASIA including Myanmar, Thailand, Malaysia, Singapore, Sumatra, Borneo, Java and Bali Indraneil Das First published in the United Kingdom in 2012 by Beaufoy Books n n 11 Blenheim Court, 316 Woodstock Road, Oxford OX2 7NS, England Contents www.johnbeaufoy.com 10 9 8 7 6 5 4 3 2 1 Introduction 4 Copyright © 2012 John Beaufoy Publishing Limited Copyright in text © Indraneil Das Snake Topography 4 Copyright in photographs © [to come] Dealing with Snake Bites 6 All rights reserved. No part of this publication may be reproduced, stored in a retrieval system or transmitted in any form or by any means, electronic, mechanical, photocopying, recording or otherwise, without the prior written permission of the publishers. About this Book 7 ISBN [to come] Glossary 8 Edited, designed and typeset by D & N Publishing, Baydon, Wiltshire, UK Printed and bound [to come] Species Accounts and Photographs 11 Checklist of South-East Asian Snakes 141 Dedication Nothing would have happened without the support of the folks at home: my wife, Genevieve V.A. Gee, and son, Rahul Das. To them, I dedicate this book. Further Reading 154 Acknowledgements 155 Index 157 Edited and designed by D & N Publishing, Baydon, Wiltshire, UK Printed and bound in Malaysia by Times Offset (M) Sdn. Bhd. n Introduction n n Snake Topography n INTRODUCTION Snakes form one of the major components of vertebrate fauna of South-East Asia. They feature prominently in folklore, mythology and other belief systems of the indigenous people of the region, and are of ecological and conservation value, some species supporting significant (albeit often illegal) economic activities (primarily, the snake-skin trade, but also sale of meat and other body parts that purportedly have medicinal properties). -

Emoia Impar and Emoia Cyanura)
bioRxiv preprint doi: https://doi.org/10.1101/2020.01.11.902866; this version posted January 14, 2020. The copyright holder for this preprint (which was not certified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available under aCC-BY 4.0 International license. SIZE-BASED DOMINANCE HIERARCHY IN ONE OF TWO SYMPATRIC CRYPTIC PACIFIC SKINKS (EMOIA IMPAR AND EMOIA CYANURA) MARY “MOLLY” HALLSTEN Department of Integrative Biology, University of California, Berkeley, California 94720 USA Abstract. Emoia impar and Emoia cyanura are two morphologically cryptic Pacific skinks that have different preferred thermal micro-habitats but similar geographic range and overlap. Previously individuals have been noted to display a size-based dominance hierarchy at favored basking sites, though this behavior was not specified between species. I found that only one of the two species, E. impar, naturally presents this size-based dominance hierarchy in areas of high population density. Neither species exhibit the hierarchy in low population density areas. No evidence was found to suggest that the presence of this hierarchy allows one species to exclude the other. Key words: skinks; Emoia; thermal resource partitioning; Moorea, French Polynesia; size- based dominance hierarchy INTRODUCTION Understanding how multiple species co- exist within a shared habitat is a fundamental challenge in ecology (Gause 1934). Hardin (1960) documented the competitive exclusion principle, which states that if two non- interbreeding populations occupy the same ecological niche and geographical territory, and differ in their rate of resource consumption, one species will eventually drive the other to extinction. -

Species Boundaries, Biogeography, and Intra-Archipelago Genetic Variation Within the Emoia Samoensis Species Group in the Vanuatu Archipelago and Oceania" (2008)
Louisiana State University LSU Digital Commons LSU Doctoral Dissertations Graduate School 2008 Species boundaries, biogeography, and intra- archipelago genetic variation within the Emoia samoensis species group in the Vanuatu Archipelago and Oceania Alison Madeline Hamilton Louisiana State University and Agricultural and Mechanical College, [email protected] Follow this and additional works at: https://digitalcommons.lsu.edu/gradschool_dissertations Recommended Citation Hamilton, Alison Madeline, "Species boundaries, biogeography, and intra-archipelago genetic variation within the Emoia samoensis species group in the Vanuatu Archipelago and Oceania" (2008). LSU Doctoral Dissertations. 3940. https://digitalcommons.lsu.edu/gradschool_dissertations/3940 This Dissertation is brought to you for free and open access by the Graduate School at LSU Digital Commons. It has been accepted for inclusion in LSU Doctoral Dissertations by an authorized graduate school editor of LSU Digital Commons. For more information, please [email protected]. SPECIES BOUNDARIES, BIOGEOGRAPHY, AND INTRA-ARCHIPELAGO GENETIC VARIATION WITHIN THE EMOIA SAMOENSIS SPECIES GROUP IN THE VANUATU ARCHIPELAGO AND OCEANIA A Dissertation Submitted to the Graduate Faculty of the Louisiana State University and Agricultural and Mechanical College in partial fulfillment of the requirements for the degree of Doctor of Philosophy in The Department of Biological Sciences by Alison M. Hamilton B.A., Simon’s Rock College of Bard, 1993 M.S., University of Florida, 2000 December 2008 ACKNOWLEDGMENTS I thank my graduate advisor, Dr. Christopher C. Austin, for sharing his enthusiasm for reptile diversity in Oceania with me, and for encouraging me to pursue research in Vanuatu. His knowledge of the logistics of conducting research in the Pacific has been invaluable to me during this process.