Cape Wind Project Is Drastically Reduced Due to the Minimal Shipping Traffic That Takes Place in the Vicinity of Horseshoe Shoal
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Competing to Overcharge Consumers: the Competitive Electric Supplier Market in Massachusetts
Why Competing to Overcharge Consumers: The Competitive Electric Supplier Market in Massachusetts April 2018 By Jenifer Bosco National Consumer Law Center® © Copyright 2018, National Consumer Law Center, Inc. All rights reserved. ABOUT THE AUTHORS Jenifer Bosco is a staff attorney at the National Consumer Law Center (NCLC) with a focus on energy and utility issues that affect low-income consumers. Prior to joining NCLC, she was the first director of the Office of Patient Protection at the Massachusetts Health Policy Commission. Previously, Jen advocated for low-income clients at Health Law Advocates, the Massachusetts Law Reform Institute and Merrimack Valley Legal Services. She also served as an Assistant Attorney General with the Office of the Massachusetts Attorney General and began her legal career as a law clerk for the Massachusetts Superior Court. Jen holds a J.D. from Georgetown University Law Center and a B.A. from Boston College. She is a co-author of Access to Utility Services and a contributing author to Collection Actions and Consumer Banking and Payments Law. ACKNOWLEDGEMENTS The author thanks NCLC Senior Attorney Charlie Harak and NCLC Deputy Director Carolyn Carter for their insightful reviews and comments, and Ana Girón-Vives of NCLC for her research and data analysis. Barbara Alexander generously shared her extensive knowledge of competitive supply markets and provided invaluable support and insight. The author also thanks Elizabeth Berube of Citizens for Citizens, Inc., Darlene Gallant of Lynn Economic Opportunity, Richard Berkley of the Public Utility Law Project of New York, and NCLC colleagues Olivia Wein, John Howat, Jan Kruse, Svetlana Ladan, and Cleef Milien for their invaluable assistance. -

Understanding the True Impacts of Champlain Hudson Power Express
UNDERSTANDING THE TRUE IMPACTS OF CHAMPLAIN HUDSON POWER EXPRESS Prepared by: Energyzt Advisors, LLC Prepared for: Independent Power Producers of New York January 2020 TABLE OF CONTENTS EXECUTIVE SUMMARY ................................................................................................... ES-1 1. OVERVIEW OF CHPE ............................................................................................................. 1 1.1. Evolution of CHPE ....................................................................................................... 3 1.2. 2017 Assessment of Carbon Reductions – Grossly Overstates Benefits ................... 4 1.3. 2019 Climate Mobilization Act – CHPE does not help New York State goals ......... 6 2. SOURCE OF CHPE ENERGY SUPPLY ................................................................................ 11 2.1 Quebec is short capacity and long on energy........................................................... 11 2.2 Hydro-Québec’s excess energy available for exports is limited ............................. 14 2.3 Hydro-Québec firm energy is very expensive ......................................................... 18 3. HYDRO-QUÉBEC WOULD DIVERT ENERGY TO SUPPLY CHPE ................................. 22 3.1. Quebec is interconnected with multiple markets .................................................... 23 3.2. Hydro-Québec has enough transmission capacity to sell its excess energy .......... 26 3.3. Hydro-Québec is motivated to maximize profits ................................................... -

Hydropower Imports
INCREMENTAL HYDROPOWER IMPORTS WHITEPAPER Fall 2013 New England States Committee on Electricity CONSIDERATIONS, OPTIONS, AND MARKET OVERVIEW REGARDING THE POTENTIAL TO INCREASE HYDROPOWER IMPORTS FROM EASTERN CANADIAN PROVINCES TO NEW ENGLAND New England States Committee on Electricity – Incremental Hydropower Imports Whitepaper Fall 2013 This whitepaper is provided solely as a source of information for New England state policymakers. The information provided is largely drawn from publicly available reports and other documents and should be independently verified before it is relied upon. Any views that may be expressed in or inferred from this whitepaper should not be construed as representing those of NESCOE, any NESCOE Manager, or any state agency or official. 2 New England States Committee on Electricity – Incremental Hydropower Imports Whitepaper Fall 2013 TABLE OF CONTENTS INTRODUCTION ............................................................................................................ 6 I. NEW ENGLAND’S COMPETITIVE WHOLESALE ELECTRICITY MARKETS, MECHANISMS TO SATISFY POLICY OBJECTIVES, AND CHALLENGES TO INTEGRATING STATE POLICIES IN THE WHOLESALE MARKETS ........................................................................................................................ 7 A. Electric Industry Restructuring and Generation Divestiture.............................. 7 B. Identifying Least-Cost Resources to Serve Customers and Examples of Existing Mechanisms to Achieve Public Policy Objectives...................................... -

STATUS of the PIPING PLOVER in MASSACHUSETTS by George W. Gove, Ashland
STATUS OF THE PIPING PLOVER IN MASSACHUSETTS by George W. Gove, Ashland On January 10, 1986, the Piping Plover (Charadrius melodus) was added to the U. S. Fish and Wildlife Service list of endangered and threatened species of wildlife. The entire breeding popula tion of this species in North America has been estimated at less than 2200 pairs. Piping Plovers breed in the Great Plains from southern Alberta eastward to Minnesota, the Dakotas, and Nebraska; at scattered locations around the Great Lakes; and on the Atlantic Coast from the north shore of the Gulf of St. Lawrence and the Maritimes to Virginia and the Carolines. They winter along the Atlantic and Gulf coasts from South Carolina to Texas and north ern Mexico. The U. S. Fish and Wildlife Service designated the Great Lakes population, which is down to less than twenty pairs, as "endangered," a term applied when extinction is imminent, and the Great Plains and Atlantic Coast populations as "threatened" (describing the state that is precursor to "endangered"). The decline of the Atlantic Coast population has been attributed to increasing recreational use and development of ocean beaches. In Massachusetts, the Piping Plover breeds coastally from Salis bury south and east to Cape Cod, the islands, and Westport. It is normally found in the state from mid-March through mid-September. This species makes a shallow nest, sometimes lined with fragments of shells, with pebbles, or wrack, along ocean beaches and filled- in areas near inlets and bays. The normal clutch of pale, sand- colored, speckled eggs is four. Incubation is underway by mid- May in Massachusetts. -

Annual Report of the Board of Harbor Commissioners, 1877 and 1878
: PUBLIC DOCUMENT .No. 33. ANNUAL REPORT OF THE Board of Harbor Commissioners FOR THE YEAR 18 7 7. BOSTON ftanb, &berg, & £0., printers to tfje CommcntntaltJ, 117 Franklin Street. 1878. ; <£ommontocaltl) of Jllaesacfjusetta HARBOR COMMISSIONERS' REPORT. To the Honorable the Senate and the Home of Representatives of the Common- wealth of Massachusetts. The Board of Harbor Commissioners, in accordance with the provisions of law, respectfully submit their annual Report. By the operation of chapter 213 of the Acts of 1877, the number of the Board was reduced from five to three members. The members of the present Board, having been appointed under the provisions of said Act, were qualified, and entered upon their duties on the second day of July. The engineers employed by the former Board were continued, and the same relations to the United States Advisory Council established. It was sought to make no change in the course of proceeding, and no interruption of works in progress or matters pending and the present Report embraces the business of the entire year. South Boston Flats. The Board is happy to announce the substantial completion of the contracts with Messrs. Clapp & Ballou for the enclosure and filling of what has been known as the Twenty-jive-acre 'piece of South Boston Flats, near the junction of the Fort Point and Main Channels, which the Commonwealth by legis- lation of 1867 (chapter 354) undertook to reclaim with mate- rial taken for the most part from the bottom of the harbor. In previous reports, more especially the tenth of the series, the twofold purpose of this work, which contemplated a har- 4 HARBOR COMMISSIONERS' REPORT. -

Docket No. EC11-35-000 Northeast Utilities
136 FERC ¶ 61,016 UNITED STATES OF AMERICA FEDERAL ENERGY REGULATORY COMMISSION Before Commissioners: Jon Wellinghoff, Chairman; Marc Spitzer, Philip D. Moeller, John R. Norris, and Cheryl A. LaFleur. NSTAR Docket No. EC11-35-000 Northeast Utilities ORDER AUTHORIZING MERGER AND DISPOSITION OF JURISDICTIONAL FACILTIES (Issued July 6, 2011) 1. On January 7, 2011, NSTAR and Northeast Utilities (collectively, Applicants) filed pursuant to sections 203(a)(1) and 203(a)(2) of the Federal Power Act (FPA)1 and Part 33 of the Commission’s regulations a joint application for authorization of a proposed transaction by which NSTAR will become a wholly-owned subsidiary of Northeast Utilities (Proposed Transaction). The Commission has reviewed the application under the Commission’s Merger Policy Statement.2 As discussed below, we will authorize the Proposed Transaction as consistent with the public interest. 1 16 U.S.C. § 824b (2006). 2 See Inquiry Concerning the Commission’s Merger Policy Under the Federal Power Act: Policy Statement, Order No. 592, FERC Stats. & Regs. ¶ 31,044 (1996), reconsideration denied, Order No. 592-A, 79 FERC ¶ 61,321 (1997) (Merger Policy Statement). See also FPA Section 203 Supplemental Policy Statement, 72 Fed. Reg. 42,277 (Aug. 2, 2007), FERC Stats. & Regs. ¶ 31,253 (2007) (Supplemental Policy Statement). See also Revised Filing Requirements Under Part 33 of the Commission’s Regulations, Order No. 642, FERC Stats. & Regs. ¶ 31,111 (2000), order on reh’g, Order No. 642-A, 94 FERC ¶ 61,289 (2001). See also Transactions Subject to FPA Section 203, Order No. 669, FERC Stats. & Regs. ¶ 31,200 (2005), order on reh’g, Order No. -

Land Protection Plan
Appendix G S. Maslowski/USFWS Least tern Land Protection Plan ■ Introduction and Purpose ■ Project Description ■ Refuge Purpose ■ Status of Resources to be Protected ■ Threats to the Resource ■ Action and Objectives ■ Protection Options ■ Land Protection Methods ■ Service Land Protection Policy ■ Funding for Fee or Easement Purchase ■ Coordination ■ Socioeconomic and Cultural Impacts ■ Literature Cited Land Protection Plan Appendix G. Land Protection Plan G-1 Project Description Introduction and Purpose This land protection plan (LPP) provides detailed information about the U.S. Fish and Wildlife Service’s (Service, we, our) proposal to expand Nantucket National Wildlife Refuge (Nantucket NWR, refuge) on Nantucket, Massachusetts. This LPP identifies the proposed land protection boundary for the Nantucket NWR. Working with numerous partners, we delineated 2,036 acres of biologically significant land on the island of Nantucket. These acres are encompassed by the recommended acquisition boundary established in alternative B of the Nantucket NWR Draft Comprehensive Conservation Plan and Environmental Assessment (draft CCP/EA). We plan to protect these lands through transfers at no cost, fee-title acquisition, conservation easements, and management agreements. Of the total acreage, we recommend acquiring 206 acres in fee title through transfers at no cost, 17 acres in fee title through purchase, 1,117 acres in conservation easements, and 696 acres through management agreement. The purposes of this LPP are to: 1) Provide landowners and the public with an outline of Service policies, priorities, and protection methods for land in the project area. Amanda Boyd/USFWS Nantucket National Wildlife Refuge 2) Assist landowners in determining whether their property lies within the proposed acquisition boundary. -

Bird Observer VOLUME 37, NUMBER 5 OCTOBER 2009 HOT BIRDS
Bird Observer VOLUME 37, NUMBER 5 OCTOBER 2009 HOT BIRDS Vern Laux, Peter Trimble, and Eddie Laux were out cruising off Nantucket when they sighted this extremely cooperative White-faced Storm Petrel (left). Peter Trimble took this fabulous photograph. Among the avian stars of the BBC Extreme Pelagic trip in early September 2009 were this Band-rumped Storm-Petrel (left) and Bridled Tern (right), both photographed by Jeremiah Trimble. On September 29, Pat Ryder told Mark Faherty about “something weird” at Wellfleet Bay Wildlife Sanctuary. Mark identified it as a Fork-tailed Flycatcher (left) and took this great photograph. Matt Garvey, Marshall Iliff, and Jeremiah Trimble were stunned to discover a Brown-chested Martin (right) at the Cumberland Farm Fields in Halifax/Middleboro on October 12, 2009. CONTENTS FALL MIGRATION HOTSPOTS IN MASSACHUSETTS: EMPHASIS ON SPARROWS Bird Observer Staff 265 FIFTY YEARS OF BIRDING: AN INTERVIEW WITH MARGARET ARGUE (reprinted from Bird Observer 21(1): 5-14) Martha Steele 277 SECOND REPORT OF THE MAINE BIRD RECORDS COMMITTEE (2007) William Sheehan, Secretary, and Peter Vickery, Chairman 287 THIRD REPORT OF THE MAINE BIRD RECORDS COMMITTEE (2008) William Sheehan, Secretary, and Peter Vickery, Chairman 290 ONA STREET WITH NO NAME John Nelson 295 ABOUT BOOKS A Modern Day Melampus Mark Lynch 298 BIRD SIGHTINGS May/June 2009 305 ABOUT THE COVER: Short-billed Dowitcher 322 ABOUT THE COVER ARTIST: Barry Van Dusen 324 ATA GLANCE Wayne R. Petersen 325 Editor’s Note: Margaret Argue died on Saturday, October 3, 2009, just two months short of her 102nd birthday. Margaret started birding with her husband Arthur in 1941 and was birding very actively right up into her nineties, when a fall curtailed her driving. -
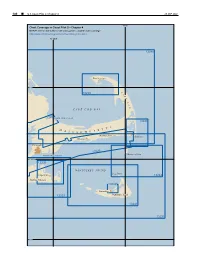
Outer Cape Cod and Nantucket Sound
186 ¢ U.S. Coast Pilot 2, Chapter 4 26 SEP 2021 70°W Chart Coverage in Coast Pilot 2—Chapter 4 NOAA’s Online Interactive Chart Catalog has complete chart coverage http://www.charts.noaa.gov/InteractiveCatalog/nrnc.shtml 70°30'W 13246 Provincetown 42°N C 13249 A P E C O D CAPE COD BAY 13229 CAPE COD CANAL 13248 T S M E T A S S A C H U S Harwich Port Chatham Hyannis Falmouth 13229 Monomoy Point VINEYARD SOUND 41°30'N 13238 NANTUCKET SOUND Great Point Edgartown 13244 Martha’s Vineyard 13242 Nantucket 13233 Nantucket Island 13241 13237 41°N 26 SEP 2021 U.S. Coast Pilot 2, Chapter 4 ¢ 187 Outer Cape Cod and Nantucket Sound (1) This chapter describes the outer shore of Cape Cod rapidly, the strength of flood or ebb occurring about 2 and Nantucket Sound including Nantucket Island and the hours later off Nauset Beach Light than off Chatham southern and eastern shores of Martha’s Vineyard. Also Light. described are Nantucket Harbor, Edgartown Harbor and (11) the other numerous fishing and yachting centers along the North Atlantic right whales southern shore of Cape Cod bordering Nantucket Sound. (12) Federally designated critical habitat for the (2) endangered North Atlantic right whale lies within Cape COLREGS Demarcation Lines Cod Bay (See 50 CFR 226.101 and 226.203, chapter 2, (3) The lines established for this part of the coast are for habitat boundary). It is illegal to approach closer than described in 33 CFR 80.135 and 80.145, chapter 2. -

Retail Competition in Electricity
33% RETAIL COMPETITION IN ELECTRICITY WHAT HAVE WE LEARNED IN 20 YEARS? July 23, 2019 Authored by: Lisa M. Quilici, Danielle S. Powers, Gregg H. Therrien, Benjamin O. Davis, and Olivia A. Prieto CEADVISORS.COM © 2019 Concentric Energy Advisors, Inc. 1 Table of Contents I. Executive Summary ...................................................................................................................................................... 1 A. Regulation and Restructuring ............................................................................................................................... 1 B. Survey of U.S. States .................................................................................................................................................. 1 C. Retail Rates for Residential Customers ............................................................................................................. 3 D. The Need for a Fully Functioning Wholesale Market .................................................................................. 4 E. Resource Planning and Reliability....................................................................................................................... 4 F. Generation Divestiture and Stranded Costs .................................................................................................... 5 G. Transition in Generation Fleet .............................................................................................................................. 5 H. Innovation in the -

November 23, 1987 Scott M. Melvin Natural Heritage And
STATUS OF PIPING PLOVEES IN MASSACHUSETTS: 1987 SUMMARY November 23, 1987 Scott M. Melvin Natural Heritage and Endangered Species Program Massachusetts Division of Fisheries and Wildlife 100 Cambridge St. Boston, MA. 02202 STATUS OF PIPING PLOVERS IN MASSACHUSETTS: 1987 SUMMARY Observers reported a total of 126 breeding pairs of Piping Plovers (Charadrius melodus) from 49 sites in Massachusetts in 1987 (Table 1). The 1987 total was lower than in either of the previous 2 years - 139 pairs (1986) and 132 pairs (1985). Observer effort in 1987, measured as number of sites surveyed and intensity of census efforts at each site, was greater than in 1985 and at least comparable to that in 1986. Thus, we believe the 1987 census results reflect a real population decline. We define breeding pairs as pairs that are observed with .either a nest or unfledged chicks, or that exhibit site tenacity and evidence of pair bonding and territoriality. Single birds that are observed during the breeding season and exhibit agitated, site tenacious behavior or distraction displays when approached are generally considered indicative of a breeding pair at a site. Breeding plovers were present at coastal sites from Plum Island south to the Rhode Island border and east to Cape Cod and Nantucket. All but 10 pairs (92%) occurred south of Boston, and 66 pairs (52%) occurred at sites on Cape Cod. More specifically, 10 pairs (8%) occurred at sites on the North Shore (New Hampshire border south to Boston), 10 pairs (8%) along the South Shore (Boston south to the Cape Cod Canal), 29 pairs (23%) at lower Cape Cod sites (Cape Cod Canal east to Brewster and Harwich) , 37 pairs (29%) on the outer Cape (Chatham and Monomoy NWR north to Provincetown), 20 pairs (16%) along the west shore of Buzzards Bay (Wareham south to Rhode Island border) and 20 pairs (16%) on the outer islands (Elizabeth Islands east to Nantucket). -

Inventory of Terns, Laughing Gulls, and Black Skimmers Nesting in Massachusetts in 2016
INVENTORY OF TERNS, LAUGHING GULLS, AND BLACK SKIMMERS NESTING IN MASSACHUSETTS IN 2016 INVENTORY OF TERNS, LAUGHING GULLS, AND BLACK SKIMMERS NESTING IN MASSACHUSETTS IN 2016 A Common Tern feeding a fledgling at Ram Island, Mattapoisett, MA. © Bill Byrne/MassWildlife, 2016. Prepared by: Carolyn S. Mostello, Jennifer Longsdorf, and Amanda Veinotte Massachusetts Division of Fisheries and Wildlife February 2018 INVENTORY OF TERNS, LAUGHING GULLS, AND BLACK SKIMMERS NESTING IN MASSACHUSETTS IN 2016 Abstract Cooperators in Massachusetts surveyed approximately 209 coastal sites in 2016 for the presence of breeding Roseate Terns (Sterna dougallii), Common Terns (Sterna hirundo), Arctic Terns (Sterna paradisaea), Least Terns (Sternula antillarum), Laughing Gulls (Larus atricilla), and Black Skimmers (Rhynchops niger). Of approximately 177 sites with a history of nesting by these species, 80 sites were occupied by nesting birds. Compared to 2015 numbers, total nesting pairs of Roseate Terns (2,066 pairs) increased 7%, Least Terns (3,230 pairs) decreased 12%, and Laughing Gulls (2,751 pairs) increased 26%. Common Terns (17,175 pairs), Arctic Terns (0.5 pairs), and Black Skimmers (6 pairs) were roughly stable. Earliest and median first egg dates were 6 May/18 May for Common Terns, 12 May/27 May for Least Terns, 15 May/19 May for Roseate Terns, 20 May/20 May for Laughing Gull, and 26 May/26 May for Black Skimmer. Introduction Coastal waterbirds are a vulnerable group of species. Of Massachusetts’ four nesting tern species, the Common Tern, the Least Tern, and the Arctic Tern are listed as “Special Concern” pursuant to the Massachusetts Endangered Species Act.