Modelling Biogeochemical Controls on Planetary Habitability
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Lecture-29 (PDF)
Life in the Universe Orin Harris and Greg Anderson Department of Physics & Astronomy Northeastern Illinois University Spring 2021 c 2012-2021 G. Anderson., O. Harris Universe: Past, Present & Future – slide 1 / 95 Overview Dating Rocks Life on Earth How Did Life Arise? Life in the Solar System Life Around Other Stars Interstellar Travel SETI Review c 2012-2021 G. Anderson., O. Harris Universe: Past, Present & Future – slide 2 / 95 Dating Rocks Zircon Dating Sedimentary Grand Canyon Life on Earth How Did Life Arise? Life in the Solar System Life Around Dating Rocks Other Stars Interstellar Travel SETI Review c 2012-2021 G. Anderson., O. Harris Universe: Past, Present & Future – slide 3 / 95 Zircon Dating Zircon, (ZrSiO4), minerals incorporate trace amounts of uranium but reject lead. Naturally occuring uranium: • U-238: 99.27% • U-235: 0.72% Decay chains: • 238U −→ 206Pb, τ =4.47 Gyrs. • 235U −→ 207Pb, τ = 704 Myrs. 1956, Clair Camron Patterson dated the Canyon Diablo meteorite: τ =4.55 Gyrs. c 2012-2021 G. Anderson., O. Harris Universe: Past, Present & Future – slide 4 / 95 Dating Sedimentary Rocks • Relative ages: Deeper layers were deposited earlier • Absolute ages: Decay of radioactive isotopes old (deposited last) oldest (depositedolder first) c 2012-2021 G. Anderson., O. Harris Universe: Past, Present & Future – slide 5 / 95 Grand Canyon: Earth History from 200 million - 2 billion yrs ago. Dating Rocks Life on Earth Earth History Timeline Late Heavy Bombardment Hadean Shark Bay Stromatolites Cyanobacteria Q: Earliest Fossils? Life on Earth O2 History Q: Life on Earth How Did Life Arise? Life in the Solar System Life Around Other Stars Interstellar Travel SETI Review c 2012-2021 G. -

A Flexible Bayesian Framework for Assessing Habitability with Joint Observational and Model Constraints
A Flexible Bayesian Framework for Assessing Habitability with Joint Observational and Model Constraints Amanda R. Truitt1, Patrick A. Young2, Sara I. Walker2, Alexander Spacek1 ABSTRACT The catalog of stellar evolution tracks discussed in our previous work is meant to help characterize exoplanet host-stars of interest for follow-up observations with future missions like JWST. However, the utility of the catalog has been predicated on the assumption that we would precisely know the age of the par- ticular host-star in question; in reality, it is unlikely that we will be able to accurately estimate the age of a given system. Stellar age is relatively straight- forward to calculate for stellar clusters, but it is difficult to accurately measure the age of an individual star to high precision. Unfortunately, this is the kind of information we should consider as we attempt to constrain the long-term habit- ability potential of a given planetary system of interest. This is ultimately why we must rely on predictions of accurate stellar evolution models, as well a con- sideration of what we can observably measure (stellar mass, composition, orbital radius of an exoplanet) in order to create a statistical framework wherein we can identify the best candidate systems for follow-up characterization. In this paper we discuss a statistical approach to constrain long-term planetary habitability by evaluating the likelihood that at a given time of observation, a star would have a planet in the 2 Gy continuously habitable zone (CHZ2). Additionally, we will discuss how we can use existing observational data (i.e. -

Ices on Mercury: Chemistry of Volatiles in Permanently Cold Areas of Mercury’S North Polar Region
Icarus 281 (2017) 19–31 Contents lists available at ScienceDirect Icarus journal homepage: www.elsevier.com/locate/icarus Ices on Mercury: Chemistry of volatiles in permanently cold areas of Mercury’s north polar region ∗ M.L. Delitsky a, , D.A. Paige b, M.A. Siegler c, E.R. Harju b,f, D. Schriver b, R.E. Johnson d, P. Travnicek e a California Specialty Engineering, Pasadena, CA b Dept of Earth, Planetary and Space Sciences, University of California, Los Angeles, CA c Planetary Science Institute, Tucson, AZ d Dept of Engineering Physics, University of Virginia, Charlottesville, VA e Space Sciences Laboratory, University of California, Berkeley, CA f Pasadena City College, Pasadena, CA a r t i c l e i n f o a b s t r a c t Article history: Observations by the MESSENGER spacecraft during its flyby and orbital observations of Mercury in 2008– Received 3 January 2016 2015 indicated the presence of cold icy materials hiding in permanently-shadowed craters in Mercury’s Revised 29 July 2016 north polar region. These icy condensed volatiles are thought to be composed of water ice and frozen Accepted 2 August 2016 organics that can persist over long geologic timescales and evolve under the influence of the Mercury Available online 4 August 2016 space environment. Polar ices never see solar photons because at such high latitudes, sunlight cannot Keywords: reach over the crater rims. The craters maintain a permanently cold environment for the ices to persist. Mercury surface ices magnetospheres However, the magnetosphere will supply a beam of ions and electrons that can reach the frozen volatiles radiolysis and induce ice chemistry. -

Life Beyond Earth the Search for Habitable Worlds in the Universe
C:/ITOOLS/WMS/CUP-NEW/4181325/WORKINGFOLDER/COEN/9781107026179HTL.3D i [1–2] 27.6.2013 10:59AM Life Beyond Earth The Search for Habitable Worlds in the Universe With current missions to Mars and the Earth-like moon Titan, and many more missions planned, humankind stands on the verge of exciting progress and possible major discoveries in our quest for life in space. What is life and where can it exist? What searches are being made to identify conditions for life on other worlds? If extraterrestrial inhabited worlds are found, how can we explore them? Could humans survive beyond the Earth? In this book, two leading astrophysicists provide an engaging account of where we stand in our quest for habitable environments, in the Solar System and beyond. Starting from basic concepts, the narrative builds up scientifically, including more in-depth material as boxed additions to the main text. The authors recount fascinating recent discoveries, arising from space missions and from observations using ground-based telescopes, of possible life-related artefacts in Martian meteorites, of extrasolar planets, and of subsurface oceans on Europa, Titan and Enceladus. They also provide a forward look to exciting future missions, including the return to Venus, Mars and the Moon; further explorations of Pluto and Jupiter’s icy moons; and placing giant planet-seeking telescopes in orbit beyond Jupiter. showing how we approach the question of finding out whether the life that teems on our own planet is unique. This is an exciting, informative read for anyone interested in the search for habitable and inhabited planets, and makes an excellent primer for students keen to learn about astrobiology, habitability, planetary science and astronomy. -

Lecture 9—Is the Earth Rare?
41st Saas-Fee Course From Planets to Life 3-9 April 2011 Lecture 9—Is the Earth Rare? List of Rare Earth arguments/ Nitrogen abundance/ Frequency of large impacts/ Chaotic obliquity fluctuations J. F. Kasting The Gaia hypothesis First presented in the 1970s by James Lovelock 1979 1988 • Life itself is what stabilizes planetary environments • Corollary: A planet might need to be inhabited in order to remain habitable The Medea and Rare Earth hypotheses Peter Ward 2009 2000 Medea hypothesis: Life is harmful to the Earth! Rare Earth hypothesis: Complex life (animals, including humans) is rare in the universe Additional support for the Rare Earth hypothesis • Lenton and Watson think that complex life (animal life) is rare because it requires a series of unlikely evolutionary events – the origin of life – the origin of the genetic code – the development of oxygenic photosynthesis) – the origin of eukaryotes – the origin of sexuality 2011 Rare Earth/Gaia arguments 1. Plate tectonics is rare --We have dealt with this already. Plate tectonics is not necessarily rare, but it requires liquid water. Thus, a planet needs to be within the habitable zone. 2. Other planets may lack magnetic fields and may therefore have harmful radiation environments and be subject to loss of atmosphere – We have talked about this one also. Venus has retained its atmosphere. The atmosphere itself provides protection against cosmic rays 3. The animal habitable zone (AHZ) is smaller than the habitable zone (HZ) o – AHZ definition: Ts = 0-50 C o – HZ definition: Ts = 0-100 C. But this is wrong! For a 1-bar atmosphere like Earth, o water loss begins when Ts reaches 60 C. -

The Search for Another Earth – Part II
GENERAL ARTICLE The Search for Another Earth – Part II Sujan Sengupta In the first part, we discussed the various methods for the detection of planets outside the solar system known as the exoplanets. In this part, we will describe various kinds of exoplanets. The habitable planets discovered so far and the present status of our search for a habitable planet similar to the Earth will also be discussed. Sujan Sengupta is an 1. Introduction astrophysicist at Indian Institute of Astrophysics, Bengaluru. He works on the The first confirmed exoplanet around a solar type of star, 51 Pe- detection, characterisation 1 gasi b was discovered in 1995 using the radial velocity method. and habitability of extra-solar Subsequently, a large number of exoplanets were discovered by planets and extra-solar this method, and a few were discovered using transit and gravi- moons. tational lensing methods. Ground-based telescopes were used for these discoveries and the search region was confined to about 300 light-years from the Earth. On December 27, 2006, the European Space Agency launched 1The movement of the star a space telescope called CoRoT (Convection, Rotation and plan- towards the observer due to etary Transits) and on March 6, 2009, NASA launched another the gravitational effect of the space telescope called Kepler2 to hunt for exoplanets. Conse- planet. See Sujan Sengupta, The Search for Another Earth, quently, the search extended to about 3000 light-years. Both Resonance, Vol.21, No.7, these telescopes used the transit method in order to detect exo- pp.641–652, 2016. planets. Although Kepler’s field of view was only 105 square de- grees along the Cygnus arm of the Milky Way Galaxy, it detected a whooping 2326 exoplanets out of a total 3493 discovered till 2Kepler Telescope has a pri- date. -
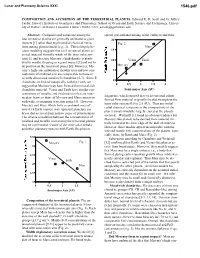
Composition and Accretion of the Terrestrial Planets
Lunar and Planetary Science XXXI 1546.pdf COMPOSITION AND ACCRETION OF THE TERRESTRIAL PLANETS. Edward R. D. Scott and G. Jeffrey Taylor, Hawai’i Institute of Geophysics and Planetology, School of Ocean and Earth Science and Technology, Univer- sity of Hawai’i at Manoa, Honolulu, Hawai’i 96822, USA; [email protected] Abstract: Compositional variations among the spread gravitational mixing of the embryos and their four terrestrial planets are generally attributed to giant 20 impacts [1] rather than to primordial chemical varia- Fig. 2 tions among planetesimals [e.g., 2]. This is largely be- Mars cause modeling suggests that each terrestrial planet ac- 15 creted material from the whole of the inner solar sys- tem [1], and because Mercury’s high density is attrib- 10 Venus Earth uted to mantle stripping in a giant impact [3] and not to its position as the innermost planet [4]. However, Mer- Mercury cury’s high concentration of metallic iron and low con- 5 centration of oxidized iron are comparable to those in recently discovered metal-rich chondrites [5-7]. Since E 0 chondrites are linked isotopically with the Earth, we 0 0.5 1 1.5 2 suggest that Mercury may have formed from metal-rich chondritic material. Venus and Earth have similar con- Semi-major Axis (AU) centrations of metallic and oxidized iron that are inter- fragments, which ensured that each terrestrial planet mediate between those of Mercury and Mars consistent formed from material originally located throughout the with wide, overlapping accretion zones [1]. However, inner solar system (0.5 to 2.5 AU). -

POSSIBLE STRUCTURE MODELS for the TRANSITING SUPER-EARTHS:KEPLER-10B and 11B
43rd Lunar and Planetary Science Conference (2012) 1290.pdf POSSIBLE STRUCTURE MODELS FOR THE TRANSITING SUPER-EARTHS:KEPLER-10b AND 11b. P. Futó1 1 Department of Physical Geography, University of West Hungary, Szombathely, Károlyi Gáspár tér, H- 9700, Hungary ([email protected]) Introduction:Up to january of 2012,10 super- The planet Kepler-11b has a large radius (1.97 R⊕) for Earths have been announced by Kepler-mission [1] its mass (4.3 M⊕),therefore this planet must have a that is designed to detect hundreds of transiting exo- spherical shell that is composed of low-density materi- planets.Kepler was launched on 6th March,2009 and the als.Considering the planet's average density,it must primary purpose of its scientific program is to search have a metallic core with different possible fractional for terrestrial-sized planets in the habitable zone of mass.Accordinghly,I have made a possible structure Solar-like stars.For the case of high number of discov- model for Kepler-11b in which the selected core mass eries we will be able to estimate the frequency of fraction is 32.59% (similarly to that of Earth) and the Earth-sized planets in our galaxy.Results of the Kepler- water ice layer has a relatively great fractional 's measurements show that the small-sized planets are volume.For case of the selected composition, the icy frequent in the spiral galaxies.A catalog of planetary surface sublimated to form a water vapor as the planet candidates,including objects with small-sized candidate moved inward the central star during its migration. -

Vol 10 No 1, Winter 2004
SearchLites Vol. 10 No. 1, Winter 2004 The Quarterly Newsletter of The SETI League, Inc. Offices: 433 Liberty Street The Date Equation PO Box 555 by David Grinspoon Little Ferry NJ 07643 USA Editor's Note: This delightful analogy on the Drake Equation is excerpted from Dr. Grinspoon's new book Lonely Planets, ISBN 0-06-018540-6, © 2003, Harper Collins Publishers, New York. Phone: (201) 641-1770 Facsimile: Say you are a single person going to a large dance party, and you would like to come away with a (201) 641-1771 date for the following weekend. Arriving in front of the house, you can hear the music pumping Email: and feel the bass rattling your gut. You are excited, but nervous as hell, so you decide to calm [email protected] yourself with some math. Before going inside, you try to calculate your chances of getting lucky. Web: You start by guessing the total number of people at the party. You notice that people are arriving www.setileague.org at a rate of three per minute. We'll call this rate of arrival R. People are leaving at roughly the President: same rate, but you realize that you can estimate the number of people inside if you know how long Richard Factor they are staying. Let's call this length of stay L. The number of people inside will be roughly R Registered Agent: times L. So, if people on average are staying for, say, one hundred minutes, there will be about Marc Arnold, Esq. three hundred inside. -

The Subsurface Habitability of Small, Icy Exomoons J
A&A 636, A50 (2020) Astronomy https://doi.org/10.1051/0004-6361/201937035 & © ESO 2020 Astrophysics The subsurface habitability of small, icy exomoons J. N. K. Y. Tjoa1,?, M. Mueller1,2,3, and F. F. S. van der Tak1,2 1 Kapteyn Astronomical Institute, University of Groningen, Landleven 12, 9747 AD Groningen, The Netherlands e-mail: [email protected] 2 SRON Netherlands Institute for Space Research, Landleven 12, 9747 AD Groningen, The Netherlands 3 Leiden Observatory, Leiden University, Niels Bohrweg 2, 2300 RA Leiden, The Netherlands Received 1 November 2019 / Accepted 8 March 2020 ABSTRACT Context. Assuming our Solar System as typical, exomoons may outnumber exoplanets. If their habitability fraction is similar, they would thus constitute the largest portion of habitable real estate in the Universe. Icy moons in our Solar System, such as Europa and Enceladus, have already been shown to possess liquid water, a prerequisite for life on Earth. Aims. We intend to investigate under what thermal and orbital circumstances small, icy moons may sustain subsurface oceans and thus be “subsurface habitable”. We pay specific attention to tidal heating, which may keep a moon liquid far beyond the conservative habitable zone. Methods. We made use of a phenomenological approach to tidal heating. We computed the orbit averaged flux from both stellar and planetary (both thermal and reflected stellar) illumination. We then calculated subsurface temperatures depending on illumination and thermal conduction to the surface through the ice shell and an insulating layer of regolith. We adopted a conduction only model, ignoring volcanism and ice shell convection as an outlet for internal heat. -

Distribution of Nitrifying Bacteria Under Fluctuating Environmental Conditions
Distribution of Nitrifying Bacteria Under Fluctuating Environmental Conditions Joseph M. Battistelli Stratford, Connecticut B.S., Rutgers, the State Universi ty of New Jersey, New Brunswick Campus, 2002 A Dissertation presented to the Graduate Faculty of the University of Virginia in Candidacy for the Degree of Doctor of Philosophy Department of Environmental Sciences University of Virginia December, 2012 ii Abstract Engineered systems that mimic tidal wetlands are ideal for point-of-source treatment of wastewater due to their low energy requirements and small physical footprint. In this study, vertical columns were constructed to mimic the flood-and-drain cycles of tidal wetland treatment systems (TWTS) in order to study how variations in the frequency and duration of flooding affect the efficiency of microbially-mediated nitrogen + removal from synthetic wastewater (containing whey protein and NH 4 ). Altering the frequency of flooding, which determines the temporal juxtaposition of aerobic and anaerobic conditions in the reactor, had a significant effect on overall nitrogen removal; + columns more frequent cycling were very efficient at converting the NH4 in the feed to - - NO 3 . At a flooding frequency of 8-cycles per day, NO 3 began to disappear from the systems in both High- and Low - N treatments. The longer flooding duration appeared to increased anaerobiosis and allowed denitrification to proceed more effectively, while allowing nitrification to proceed when oxygen was available. Analysis of depth profiles of abundance revealed distinct differences between the tidal and trickling systems abundance profiles. Overall, these results demonstrate a tight coupling of environmental conditions with the abundance of ammonium oxidizing bacteria and suggest several experimental modifications, such variable tidal cycles, could be implemented to enhance the functioning of TWTS. -

Exep Science Plan Appendix (SPA) (This Document)
ExEP Science Plan, Rev A JPL D: 1735632 Release Date: February 15, 2019 Page 1 of 61 Created By: David A. Breda Date Program TDEM System Engineer Exoplanet Exploration Program NASA/Jet Propulsion Laboratory California Institute of Technology Dr. Nick Siegler Date Program Chief Technologist Exoplanet Exploration Program NASA/Jet Propulsion Laboratory California Institute of Technology Concurred By: Dr. Gary Blackwood Date Program Manager Exoplanet Exploration Program NASA/Jet Propulsion Laboratory California Institute of Technology EXOPDr.LANET Douglas Hudgins E XPLORATION PROGRAMDate Program Scientist Exoplanet Exploration Program ScienceScience Plan Mission DirectorateAppendix NASA Headquarters Karl Stapelfeldt, Program Chief Scientist Eric Mamajek, Deputy Program Chief Scientist Exoplanet Exploration Program JPL CL#19-0790 JPL Document No: 1735632 ExEP Science Plan, Rev A JPL D: 1735632 Release Date: February 15, 2019 Page 2 of 61 Approved by: Dr. Gary Blackwood Date Program Manager, Exoplanet Exploration Program Office NASA/Jet Propulsion Laboratory Dr. Douglas Hudgins Date Program Scientist Exoplanet Exploration Program Science Mission Directorate NASA Headquarters Created by: Dr. Karl Stapelfeldt Chief Program Scientist Exoplanet Exploration Program Office NASA/Jet Propulsion Laboratory California Institute of Technology Dr. Eric Mamajek Deputy Program Chief Scientist Exoplanet Exploration Program Office NASA/Jet Propulsion Laboratory California Institute of Technology This research was carried out at the Jet Propulsion Laboratory, California Institute of Technology, under a contract with the National Aeronautics and Space Administration. © 2018 California Institute of Technology. Government sponsorship acknowledged. Exoplanet Exploration Program JPL CL#19-0790 ExEP Science Plan, Rev A JPL D: 1735632 Release Date: February 15, 2019 Page 3 of 61 Table of Contents 1.