Entomologic and Molecular Investigation Into Plasmodium Vivax
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

List of Publicly Accessible Locations Where E-Bins Are Deployed*
List of publicly accessible locations where E-Bins are deployed* *This is a working list, more locations will be added every week* Name Location Type of Bin Placed Best Denki • 1 Harbourfront Walk, Vivocity, #2-07 • 3155 Commonwealth Avenue West, The Clementi Mall, #04-46/47/48/49 • 68 Orchard Road, Plaza Singapura, #3-39 • 2 Jurong East Street 21, IMM, #3-33 • 63 Jurong West Central 3, Jurong Point, #B1-92 • 109 North Bridge Road, Funan, #3-16 3-in-1 Bin • 1 Kim Seng Promenade, Great World City, #07-01 (ICT, Bulb, Battery) • 391A Orchard Road, Ngee Ann City Tower A • 9 Bishan Place, Junction 8 Shopping Centre, #03-02 • 17 Petir Road, Hillion Mall, #B1-65 • 83 Punggol Central, Waterway Point • 311 New Upper Changi Road, Bedok Mall Cash • 135 Jurong Gateway Road 3-in-1 Bin Converters • 510 Tampines Central 1 (ICT, Bulb, Battery) Challenger • 302 Tiong Bahru Road, Tiong Bahru Plaza, #03-19 • 1 Jurong West Central 2, Jurong Point, #B1-94 • 200 Victoria Street, Bugis Junction, #03-10E • 5 Changi Business Park Central 1, Changi City Point, #01-56 3-in-1 Bin • 3 Simei Street 6, Eastpoint Mall, #B1-08 (ICT, Bulb, Battery) • 90 Hougang Avenue 10, Hougang Mall, #02-13 • 10 Paya Lebar Road, Paya Lebar Quarter (PLQ), #04-01 • 10 Tampines Central 1, Tampines 1, #04-25 City Square Mall • 180 Kitchener Road 3-in-1 Bin (ICT, Bulb, Battery) Compass One • 1 Sengkang Square, #B1-52/53 3-in-1 Bin (ICT, Bulb, Battery) Dairy Farm • Cold Storage, 5 Stadium Walk, Kallang Leisure Park, #B1-01 Group • Giant Hypermarket, 521 Tampines North Drive 2, #03-01, Singapore -

List of Licensed General Waste Disposal Facilities (Gwdfs) IMPORTANT NOTE: Please Contact the Companies for More Information
List of Licensed General Waste Disposal Facilities (GWDFs) IMPORTANT NOTE: Please contact the companies for more information. Since 1 August 2017, NEA began licensing General Waste Disposal Facilities (GWDFs). A GWDF is defined as a disposal facility which receives, stores, sorts, treats or processes general waste, and includes recycling facilities. Companies can apply for the Licence/Exemption via https://licence1.business.gov.sg. All general waste disposal facilities must obtain their licence or submit an exemption declaration by 31 July 2018. For more information on the GWDF Licence, please visit http://www.nea.gov.sg/energy-waste/waste-management/general-waste-disposal-facility/ Waste Stream Company Facility Address Contacts Ash Paper Plastic Sludge E-Waste Steel Slag Steel C&D waste C&D Refrigerant Scrap Metal Scrap Glass Waste Glass WoodWaste Textile Waste Textile Biomass Waste Biomass Return Concrete Return Used Cooking Oil Cooking Used Spent Copper Slag Spent Copper Mixed Recyclables Mixed Horticultural Waste Horticultural Tyre/RubberWaste Used CoffeeCapsules Used Refrigerant Cylinder/Tank Refrigerant Waste generated from the from generated Waste manufacture of electrical and manufactureofelectrical Industrial and Commercial Waste andCommercial Industrial 800 Super Waste Management 6 Tuas South Street 7 636892 [email protected]; Y Pte Ltd 62 Sungei Kadut Street 1 Sungei [email protected]; 85 Auto Trading Y Kadut Industrial Estate 729363 [email protected]; 21 Tuas West Avenue #03-01 A~Star Plastics Pte Ltd [email protected] -

Slides by JTC GD Calvin Chung
• Sungei Kadut Eco-District • Seletar Aerospace Park Catalysing Industrial Transformation for sustainability Mr. Calvin Chung Group Director/ Chief Environmental Officer 25 May 2021 1. National Challenges and Sustainability Targets 2 Carbon – the New Constraint for Singapore ▪ Singapore is no stranger to constraints: land scarce, lack of water. Carbon would be a new addition to the list ▪ Like Our Water Story – Singapore is embarking on another journey to address the carbon constraint Singapore carbon emission projection (Climate Action Tracker) ▪ Singapore has signed Paris Agreement and committed to 36% reduction in emission intensity below 2005 by 2030 ▪ Singapore national targets: peak emission of 65 million tons of CO2e by 2030 and to be halved by 2050 ▪ A carbon tax for industrial facilities at $5 SGD/tCo2e was implemented in 2019, to be reviewed in 2023 ▪ A 2GW target by 2030 for solar energy was unveiled Peak by 2030, in SIEW 2019 halved by 2050 4oC+ Critically Insufficient <3oC Insufficient <1.5oC Paris Agreement <4oC Highly Insufficient <2oC Compatible <<1.5oC Role Model 3 Strong National Drive Towards Sustainability – SG Green Plan 2030 • SG Green Plan - An overarching sustainability vision for Singapore, a whole-of-nation action plan to realise the vision, and a relatable communication with the public to inspire action • JTC’s ongoing sustainability effort has already been contributing to the SG Green Plan pillars. City in Nature Energy Reset Sustainable Living Green Economy Resilient Future ▪ One million more ▪ Green 80% of -

Annex I 01 September 2015
Quarantine & Inspection Group Agri-Food & Veterinary Authority of Singapore Sembawang Research Station Lorong Chencharu, Singapore 769194 PR 11.4 Annex I 01 September 2015 To: All NPPO, Traders & Forwarding Agents LIST OF AVA ACCREDITED TREATMENT PROVIDERS Please find below the list of treatment providers accredited to carry out phytosanitary treatment for solid wood packing materials under International Standards for Phytosanitary Measures (i.e. ISPM 15) and other agricultural commodities that require phytosanitary certification. These accredited treatment providers would also issue Fumigation Certificate or Heat Treatment Certificate to certify that the solid wood packaging materials or other agricultural commodities had been treated in accordance to the standard such as ISPM 15 and other treatment specifications imposed by the importing countries. 2. The list of AVA accredited treatment providers will be updated from time to time and posted in the AVA website http://www.ava.gov.sg. For any other clarifications, you can contact us at Tel No: (65) 67591-9816, (65) 6753-0657, (65) 6751-9842 or email to Mr. Eric Casiano Tulang at [email protected] or Ms. Ong Ai Khim at: [email protected]. Yours faithfully, Eric Casiano Tulang Senior Executive Manager for Director General Agri-Food & Veterinary Authority of Singapore September 2015 Page 1 of 6 Quarantine & Inspection Group Agri-Food & Veterinary Authority of Singapore Sembawang Research Station Lorong Chencharu, Singapore 769194 LIST OF AVA ACCREDITED PEST CONTROL AGENCIES TYPE OF TREATMENT: HEAT TREATMENT (HT) TPS CONTACT S/N NAME OF PEST CONTROL AGENCY ADDRESS EFFECTIVE DATE CODE NUMBER 36 Sungei Kadut Street 2 Tel: 62696265 01 HOCK HUAT SAW MILL CO. -

Submerged Outlet Drain Once in Three Month Desilting and Flushing
Submerged Outlet Drain Once in Three Month Desilting and Flushing Drain S/No Location Frequency Length 4.5m wide U-drain from the culvert at Jurong Road near Track 22 to the 1 outlet of the culvert at PIE (including the culverts across Jurong Road and 65 1st week of the month across PIE) 9m/12m wide Sungei Jurong subsidiary drain running along PIE from L/P 2 380 1st week of the month 606 to L/P 586 1.5m wide U-drain/covered drain from the junction of Yuan Ching Road/ 3 790 2nd week of the month Jalan Ahmad 13m wide U-drain from Jurong West Street 65 to Major Drain MJ 14 near 4 450 2nd week of the month Blk 664A 15m wide Sg Jurong subsidiary drain from L/P 586 at PIE to Sg Jurong 5 950 2nd week of the month including one 6 10m wide Sg Jurong subsidiary drain from L/P 534 at PIE to Sg Jurong 1180 3rd week of the month Along Boon Lay Way opposite Jurong West Street 61 to Jalan Boon Lay 7 1300 4th week of the month and at Enterprise Road 8 Blk 664A Jurong West Street 64 to MJ13 650 4th week of the month 9 Sungei Lanchar (From Jalan Boon Lay to Jurong Lake 1,600 5th week of the month 10 Sungei Jurong (From Ayer Rajah Expressway to Jurong Lake) 1070 6th week of the month 3.0m wide Outlet drain from culvert at Teban Garden Road running along 11 Jurong Town Hall Road and West Coast Road to Sg Pandan opposite Block 650 7th week of the month 408 30.0m wide Sg Pandan from the branch connection to the downstream Boon 12 350 7th week of the month Lay Way to Sg Ulu Pandan 13 Sungei Pandan (from confluence of Sg Pandan to West Coast Road) 600 7th week -

List-Of-Bin-Locations-1-1.Pdf
List of publicly accessible locations where E-Bins are deployed* *This is a working list, more locations will be added every week* Name Location Type of Bin Placed Ace The Place CC • 120 Woodlands Ave 1 3-in-1 Bin (ICT, Bulb, Battery) Apple • 2 Bayfront Avenue, B2-06, MBS • 270 Orchard Rd Battery and Bulb Bin • 78 Airport Blvd, Jewel Airport Ang Mo Kio CC • Ang Mo Kio Avenue 1 3-in-1 Bin (ICT, Bulb, Battery) Best Denki • 1 Harbourfront Walk, Vivocity, #2-07 • 3155 Commonwealth Avenue West, The Clementi Mall, #04- 46/47/48/49 • 68 Orchard Road, Plaza Singapura, #3-39 • 2 Jurong East Street 21, IMM, #3-33 • 63 Jurong West Central 3, Jurong Point, #B1-92 • 109 North Bridge Road, Funan, #3-16 3-in-1 Bin • 1 Kim Seng Promenade, Great World City, #07-01 (ICT, Bulb, Battery) • 391A Orchard Road, Ngee Ann City Tower A • 9 Bishan Place, Junction 8 Shopping Centre, #03-02 • 17 Petir Road, Hillion Mall, #B1-65 • 83 Punggol Central, Waterway Point • 311 New Upper Changi Road, Bedok Mall • 80 Marine Parade Road #03 - 29 / 30 Parkway Parade Complex Bugis Junction • 230 Victoria Street 3-in-1 Bin Towers (ICT, Bulb, Battery) Bukit Merah CC • 4000 Jalan Bukit Merah 3-in-1 Bin (ICT, Bulb, Battery) Bukit Panjang CC • 8 Pending Rd 3-in-1 Bin (ICT, Bulb, Battery) Bukit Timah Plaza • 1 Jalan Anak Bukit 3-in-1 Bin (ICT, Bulb, Battery) Cash Converters • 135 Jurong Gateway Road • 510 Tampines Central 1 3-in-1 Bin • Lor 4 Toa Payoh, Blk 192, #01-674 (ICT, Bulb, Battery) • Ang Mo Kio Ave 8, Blk 710A, #01-2625 Causeway Point • 1 Woodlands Square 3-in-1 Bin (ICT, -
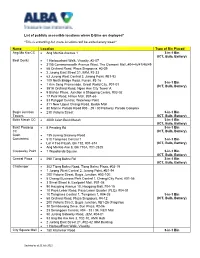
List of Publicly Accessible Locations Where E-Bins Are Deployed*
List of publicly accessible locations where E-Bins are deployed* *This is a working list, more locations will be added every week* Name Location Type of Bin Placed Ang Mo Kio CC • Ang Mo Kio Avenue 1 3-in-1 Bin (ICT, Bulb, Battery) Best Denki • 1 Harbourfront Walk, Vivocity, #2-07 • 3155 Commonwealth Avenue West, The Clementi Mall, #04-46/47/48/49 • 68 Orchard Road, Plaza Singapura, #3-39 • 2 Jurong East Street 21, IMM, #3-33 • 63 Jurong West Central 3, Jurong Point, #B1-92 • 109 North Bridge Road, Funan, #3-16 3-in-1 Bin • 1 Kim Seng Promenade, Great World City, #07-01 (ICT, Bulb, Battery) • 391A Orchard Road, Ngee Ann City Tower A • 9 Bishan Place, Junction 8 Shopping Centre, #03-02 • 17 Petir Road, Hillion Mall, #B1-65 • 83 Punggol Central, Waterway Point • 311 New Upper Changi Road, Bedok Mall • 80 Marine Parade Road #03 - 29 / 30 Parkway Parade Complex Bugis Junction • 230 Victoria Street 3-in-1 Bin Towers (ICT, Bulb, Battery) Bukit Merah CC • 4000 Jalan Bukit Merah 3-in-1 Bin (ICT, Bulb, Battery) Bukit Panjang • 8 Pending Rd 3-in-1 Bin CC (ICT, Bulb, Battery) Cash • 135 Jurong Gateway Road Converters • 510 Tampines Central 1 3-in-1 Bin • Lor 4 Toa Payoh, Blk 192, #01-674 (ICT, Bulb, Battery) • Ang Mo Kio Ave 8, Blk 710A, #01-2625 Causeway Point • 1 Woodlands Square 3-in-1 Bin (ICT, Bulb, Battery) Central Plaza • 298 Tiong Bahru Rd 3-in-1 Bin (ICT, Bulb, Battery) Challenger • 302 Tiong Bahru Road, Tiong Bahru Plaza, #03-19 • 1 Jurong West Central 2, Jurong Point, #B1-94 • 200 Victoria Street, Bugis Junction, #03-10E • 5 Changi Business -

ANNEX a OUR ECONOMIC GATEWAYS the Economic
ANNEX A OUR ECONOMIC GATEWAYS The economic gateways in the east, west and north of Singapore bring together key employment areas and transport infrastructure (e.g. airport, RTS link, port) that serve as a portal between Singapore and the rest of the world. The gateways offer strong potential for new synergies, and diversify and deepen our air, sea, and rail linkages to external markets. Eastern Gateway Changi Region is set to be a vibrant and thriving economic hub, hosting businesses and industries that leverage the connectivity of Changi Airport. Business gateway to the world Changi Region comprises two key growth nodes: Changi Aviation Park to the north, which includes the existing air cargo cluster at Changi Airfreight Centre (CAC) and Aviation Logistics Park of Singapore (ALPS) as well as the future Changi East Industrial Zone (CEIZ). Together, they cater to new aviation-related sectors and businesses that rely on air connectivity to the world. Changi City to the south, which is envisioned as an innovative lifestyle business cluster comprising the Changi Business Park, Singapore University of Technology and Design, as well as the future Changi East Urban District located at the doorstep of Changi Airport Terminal 5. Attractive lifestyle destination Beyond an employment node, Changi Region will have a diverse range of recreation and tourism attractions–from the rich, rustic heritage of Changi Point to the lifestyle and entertainment options within Jewel Changi Airport—connected by the Round Island Route and wider cycling network. In the longer term, a waterfront district to the south could be developed to offer exciting new recreational and tourism possibilities. -

Approved-Covid-19-Pcr-Swab-Provider
Ministry of Health List of Approved Offsite Providers for Polymerase Chain Reaction (PCR) Tests for COVID-19 List updated as at 08 January 2021. Service Provider Name of Location Address Service Provided Partnering Lab ST Engineering Marine 16 Benoi Road S(629889) Parkway Laboratory Services Ltd Ally Health (in Offsite PCR Swab and Quest Laboratories Pte Ltd partnership with Jaga- 432A Bukit Batok West Avenue 8, Bukit Batok North N4 Serology Q Squared Solutions Laboratory Me) S(651432) C882 6A Raeburn Park, S(088703) National Public Health Laboratory Singapore Salvage 23 Shipyard Road Singapore Engineers Pte Ltd 628129 PSA Multi-Purpose 33 Harbour Drive, Singapore Pasir Panjang Terminal 117606 1 Brani Terminal Avenue, PSA Brani Terminal Singapore 098680 37 Jurong Port Road, Singapore Jurong Port 619110 460 Alexandra Road, Singapore CITYMED HEALTH PSA Keppel Terminal Offsite PCR Swab and Home Team Science & ASSOCIATES PTE LTD 119963 Serology Technology Agency, HTX Tuas Mega Yard 80 Tuas S Blvd, Singapore 636991 Sembcorp Sembcorp Marine 60 Admiralty Rd W, Singapore Admiralty Yard 759956 75 Deptford Rd, Singapore Sembawang Wharves 759657 St Engineering Marine 7 Benoi Rd, Singapore 629882 Page 1 of 82 St Marine Tuas 60 Tuas Rd, Singapore 638501 Shipyard Keppel Shipyard - Gul 55 Gul Rd, Singapore 629353 51 Pioneer Sector 1, Singapore Keppel Shipyard - Tuas 628437 Keppel Shipyad – 15 Benoi Rd, Singapore 629888 Benoi Loyang Offshore 25 Loyang Crescent, Avenue 1 Supply Base Singapore 508988 Pax Ocean Shipyard 33 Tuas Cres, Singapore 638722 DSTAC02 -

JTC Launchpad @ One-North
ULI Summit Talk Innovative Urban Planning Mr David Tan Assistant Chief Executive Officer 6 June 2017 1 What We Do Our Strategy Map MISSION To develop industrial infrastructure to catalyse the growth of new industries and transform existing enterprises KEY FOCUS AREAS Master Develop Next Generation Estates • Plan, develop & manage mixed-use estates Develop Innovative & Flexible Spaces • Act as place maker to enhance vibrancy and connectivity of public spaces Optimise Land Use Drive Innovation & Productivity 2 Master Develop Next Generation Estates Plan, Develop & Manage Mixed-Use Estates A Master Developer goes beyond providing infrastructure Integrate mixed Promote the uses for right mix of work-live- industries play-learn Plan sustainable district-level Placemaking Test-bed shared to build services community & solutions enhance as living vibrancy lab 3 Integrate Mixed-uses for Work-Live-Play-Learn WORK LIVE PLAY LEARN Industrial Residential Recreational Educational R&D Retail Office Community Industrial & Lifestyle Office Offerings At Buildings Easy Access To Industry- Good Range of Transport Academia Amenities Nodes & Public Collaboration Residential Spaces Neighbourhoods Educational Landscaped Public / Community Programmes Recreational Spaces for Place Corridors Making Events Plan Sustainable District-level Shared Services • Buildings would be integrated by district-wide shared services to enhance operational efficiency, productivity and competitiveness Pneumatic Waste Conveyance System District Cooling System Integrated Facility Management -

(5.) Name of Dormitory (6.)Job Sector (8.) Company Name Location Address
Myanmar electors in Foreign Workers Dormitories No (1.) Name (5.) Name of Dormitory (6.)Job Sector (8.) Company Name Location Address Mun Siong Engineering 1 MOE HLAING YUNG. KUANG Process Taman Jurong 35 TUAS ROAD Singapore 638496 Limited Modern building 2 Zaw Zaw Naing Woodlands LodgeOne Construction Woodlands 45 LOYANG DRIVE LOYANG INDUSTRIAL ESTATE Singapore 508954 materials Pte Ltd CHUNG SHAN 3 Nyi Nyi Naing WOODLANDS LODGE ONE Construction Woodlands 211 Woodlands Ave 9, Singapore 738960 PLASTICS PTE,LTD. 4 Shine Aung Woodlands dormitory Construction AVon Industeries Pte Ltd Woodlands 2 Woodlands Sector 1, #01-01 Spectrum I, Singapore 757740 5 Pyae Sone Oo Woodlands Dormitory Construction Million Build Pte Ltd Woodlands 9 Yishun Industrial Street 1 #05-61/62 North Spring BizHub Singapore, 768163 6 Than Naing Oo Woodlands dormitory Construction Million Build PteLtd Woodlands 9 Yishun Industrial Street 1 #05-61/62 North Spring BizHub Singapore, 768163 7 Aung myo thu Woodlands dormitory Construction Million built pte ltd Woodlands 9 Yishun Industrial Street 1 #05-61/62 North Spring BizHub Singapore, 768163 8 Tay Zar Oo Woodlands Dormitory Construction Million Build Pte Ltd Woodlands 9 Yishun Industrial Street 1 #05-61/62 North Spring BizHub Singapore, 768163 9 Nay Lin Kyaw Woodlands Dormitory Construction CM METAL PTE. LTD Woodlands 22 WOODLANDS LINK #04-45 Singapore 738734 MODERN BUILDING 10 MIN CHAN AUNG WOODLANDS Construction Woodlands 45 LOYANG DRIVE LOYANG INDUSTRIAL ESTATE Singapore 508954 MATERIALS 11 Zaw linn oo Woodland lodge -

Ministry of Health List of Approved Offsite Providers for Polymerase Chain Reaction (PCR) Tests for COVID-19
Ministry of Health List of Approved Offsite Providers for Polymerase Chain Reaction (PCR) Tests for COVID-19 List updated as at 13 August 2021. S/N Service Provider Name of Location Address Service Partnering Lab Provided 1 Acumen Diagnostics Former Siglap Secondary School 10 Pasir Ris Drive 10, Singapore 519385 Offsite PCR Acumen Pte. Ltd K.H. Land Pte Ltd. The Antares @ Mattar Road Swab and Diagnostics Pte. Serology Ltd. Keong Hong Construction Pte Ltd National Skin Centre @ 1 Mandalay Road Keong Hong Construction Pte Ltd Sky Everton @ 42 Everton Road The Antares 23 Mattar Road, Singapore 387730 National Skin Centre 1 Mandalay Road, Singapore 308205 Sky Everton 50 Everton Road, Singapore 089396 2 ACUMED MEDICAL Shangri-La Hotel 22 Orange Grove Rd, Singapore 258350 Offsite PCR Parkway GROUP PEC Ltd 20 Benoi Lane Singapore 627810 Swab and Laboratory Services Serology Ltd LC&T Builder (1971) Pte Ltd 172A Sengkang East Drive Singapore 541172 Dyna-Mac Engineering Services Pte 59 Gul Road Singapore 629354 Ltd Jurong Fishery Port Fishery Port Road Singapore 619742 Senoko Fishery Port 31 Attap Valley Road Singapore 759908 Franklin Offshore International Pte 11 Pandan Road Singapore 609259 Ltd CFE Engineers Pte Ltd 10 Pioneer Sector Singapore 628444 Syscon Private Limited 30 Tuas Bay Drive Singapore 637548 3 Ally Health ST Engineering Marine 16 Benoi Road S(629889) Offsite PCR Parkway Bukit Batok North N4 432A Bukit Batok West Avenue 8, S(651432) Swab and Laboratory Services Serology Ltd C882 6A Raeburn Park, S(088703) Quest Laboratories CSC@Tessensohn