Language and Modern Human Origins
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

The Evolutionary History of the Human Face
This is a repository copy of The evolutionary history of the human face. White Rose Research Online URL for this paper: https://eprints.whiterose.ac.uk/145560/ Version: Accepted Version Article: Lacruz, Rodrigo S, Stringer, Chris B, Kimbel, William H et al. (5 more authors) (2019) The evolutionary history of the human face. Nature Ecology and Evolution. pp. 726-736. ISSN 2397-334X https://doi.org/10.1038/s41559-019-0865-7 Reuse Items deposited in White Rose Research Online are protected by copyright, with all rights reserved unless indicated otherwise. They may be downloaded and/or printed for private study, or other acts as permitted by national copyright laws. The publisher or other rights holders may allow further reproduction and re-use of the full text version. This is indicated by the licence information on the White Rose Research Online record for the item. Takedown If you consider content in White Rose Research Online to be in breach of UK law, please notify us by emailing [email protected] including the URL of the record and the reason for the withdrawal request. [email protected] https://eprints.whiterose.ac.uk/ THE EVOLUTIONARY HISTORY OF THE HUMAN FACE Rodrigo S. Lacruz1*, Chris B. Stringer2, William H. Kimbel3, Bernard Wood4, Katerina Harvati5, Paul O’Higgins6, Timothy G. Bromage7, Juan-Luis Arsuaga8 1* Department of Basic Science and Craniofacial Biology, New York University College of Dentistry; and NYCEP, New York, USA. 2 Department of Earth Sciences, Natural History Museum, London, UK 3 Institute of Human Origins and School of Human Evolution and Social Change, Arizona State University, Tempe, AZ. -

Two Modern Compositional Approaches
Musical Aesthetics of the Natural World: Two Modern Compositional Approaches Eli Stine and Christopher Luna-Mega University of Virginia, McIntire Department of Music Throughout recorded human history, experiences and observations of the natural world have inspired the arts. Within the sonic arts, evocations of nature permeate a wide variety of acoustic and electronic composition strategies. These strategies artistically investigate diverse attributes of nature: tranquility, turbulence, abundance, scarcity, complexity, and purity, to name but a few. Within the 20th century, new technologies to understand these attributes, including media recording and scientific analysis, were developed. These technologies allow music composi- tion strategies to go beyond mere evocation and to allow for the construction of musical works that engage explicit models of nature (what has been called ‘biologically inspired music’). This paper explores two such deployments of these ‘natural sound models’ within music and music generation systems created by the authors: an electroacoustic composition using data derived from multi-channel recordings of forest insects (Luna-Mega) and an electronic music generation system that extracts musical events from the different layers of natural soundscapes, in particular oyster reef soundscapes (Stine). Together these works engage a diverse array of extra-musical disciplines: environmental science, acoustic ecology, entomology, and computer science. The works are contextualized with a brief history of natural sound models from pre- antiquity to the present in addition to reflections on the uses of technology within these projects and the potential experiences of audiences listening to these works. “Great art picks up where nature ends.” - Mark Chagall and aesthetics towards a more quantitative awareness of how The natural world has been a focal point for the arts since the natural world functions and evolves is present within a the Upper Paleolithic. -
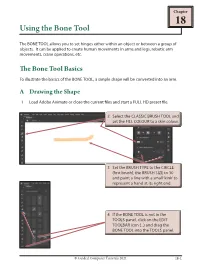
Using the Bone Tool
Chapter 18 Using the Bone Tool The BONE TOOL allows you to set hinges either within an object or between a group of objects. It can be applied to create human movements in arms and legs, robotic arm movements, crane operations, etc. The Bone Tool Basics To illustrate the basics of the BONE TOOL, a simple shape will be converted into an arm. A Drawing the Shape 1 Load Adobe Animate or close the current files and start a FULL HD preset file. © Guided Computer Tutorials 2021 18-1 Learning Adobe Animate CC B Applying the Bone Tool 1 Press CTRL+ or COMMAND+ to zoom the view to 200%. 3 When you release the mouse button the first bone is created. NOTE: This first section will represent a bone from the shoulder to the elbow. 5 Release the mouse button to create the second bone. NOTE: This second section will represent a bone from the elbow to the wrist. 18-2 © Guided Computer Tutorials 2021 Using the Bone Tool 18 NOTE: All the bone sections are moved into the ARMATURE layer. C Using the Bone Links The bone sections have set a rotation joint at the left of the shape (red diamond shape) and hinges (or joints) at the centre and near the right of the shape. 1 Press CTRL- or COMMAND- to return the view to 100%. NOTE: When the mouse pointer is over a joint or bone that can be moved, a bone symbol is added to the pointer. © Guided Computer Tutorials 2021 18-3 Learning Adobe Animate CC D The Pin Option The PIN option allows you to fix the position of a bone and prevent it from moving. -

Adobe Animate Cc Classroom in a Book (2018 Release) 301
NATURAL AND CHARACTER 9 ANIMATION Lesson Overview In this lesson, you’ll learn how to do the following: • Use the Bone tool to build armatures (skeletons) of movie clips • Use the Bone tool to build armatures of shapes • Animate natural motion of armatures using inverse kinematics • Constrain and pin the armature joints • Edit the position of armature bones and joints • Refine shape deformations with the Bind tool • Simulate physics with the Spring feature • Adjust the speed setting to add a sense of weight to armatures This lesson will take about one and a half hours to complete. Please log in to your account on peachpit.com to download the lesson files for this chap- ter, or go to the Getting Started section at the beginning of this book and follow the instructions under “Accessing the Lesson Files and Web Edition.” 298 From the Library of Alvaro Alvarez You can easily create complex and natural motion with articulations—joints between linked objects and within shapes—by using the Bone tool for animation in a process called inverse kinematics . 299 From the Library of Alvaro Alvarez Getting Started You’ll start the lesson by viewing the animated walking monkey that you’ll create as you learn about natural motion in Adobe Animate CC. 1 Double-click the 09End.html file in the Lesson09/09End folder to play the animation. The animation depicts a cartoon monkey walking in an endless cycle with a scrolling motion in the background. His arms and legs swing naturally, and his tail curls and unfurls naturally and smoothly. -

Language Evolution to Revolution: from a Slowly Developing Finite Communication System with Many Words to Infinite Modern Language
bioRxiv preprint doi: https://doi.org/10.1101/166520; this version posted July 20, 2017. The copyright holder for this preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. Language evolution to revolution: from a slowly developing finite communication system with many words to infinite modern language Andrey Vyshedskiy1,2* 1Boston University, Boston, USA 2ImagiRation LLC, Boston, MA, USA Keywords: Language evolution, hominin evolution, human evolution, recursive language, flexible syntax, human language, syntactic language, modern language, Cognitive revolution, Great Leap Forward, Upper Paleolithic Revolution, Neanderthal language Abstract There is overwhelming archeological and genetic evidence that modern speech apparatus was acquired by hominins by 600,000 years ago. There is also widespread agreement that modern syntactic language arose with behavioral modernity around 100,000 years ago. We attempted to answer two crucial questions: (1) how different was the communication system of hominins before acquisition of modern language and (2) what triggered the acquisition of modern language 100,000 years ago. We conclude that the communication system of hominins prior to 100,000 years ago was finite and not- recursive. It may have had thousands of words but was lacking flexible syntax, spatial prepositions, verb tenses, and other features that enable modern human language to communicate an infinite number of ideas. We argue that a synergistic confluence of a genetic mutation that dramatically slowed down the prefrontal cortex (PFC) development in monozygotic twins and their spontaneous invention of spatial prepositions 100,000 years ago resulted in acquisition of PFC-driven constructive imagination (mental synthesis) and converted the finite communication system of their ancestors into infinite modern language. -

Upper Pleistocene Human Remains from Vindija Cave, Croatia, Yugoslavia
AMERICAN JOURNAL OF PHYSICAL ANTHROPOLOGY 54~499-545(1981) Upper Pleistocene Human Remains From Vindija Cave, Croatia, Yugoslavia MILFORD H. WOLPOFF, FRED H. SMITH, MIRKO MALEZ, JAKOV RADOVCIC, AND DARKO RUKAVINA Department of Anthropology, University of Michigan, Ann Arbor, Michigan 48109 (MH.W.1,Department of Anthropology, University of Tennessee, Kmxuille, Tennessee 37916 (FHS.),and Institute for Paleontology and Quaternary Geology, Yugoslav Academy of Sciences and Arts, 41000 Zagreb, Yugoslavia (M.M., J.R., D.R.) KEY WORDS Vindija, Neandertal, South Central Europe, Modern Homo sapiens origin, Evolution ABSTRACT Human remains excavated from Vindija cave include a large although fragmentary sample of late Mousterian-associated specimens and a few additional individuals from the overlying early Upper Paleolithic levels. The Mousterian-associated sample is similar to European Neandertals from other regions. Compared with earlier Neandertals from south central Europe, this sam- ple evinces evolutionary trends in the direction of Upper Paleolithic Europeans. Compared with the western European Neandertals, the same trends can be demon- strated, although the magnitude of difference is less, and there is a potential for confusing temporal with regional sources of variation. The early Upper Paleo- lithic-associated sample cannot be distinguished from the Mousterian-associated hominids. We believe that this site provides support for Hrdlicka’s “Neandertal phase” of human evolution, as it was originally applied in Europe. The Pannonian Basin and surrounding val- the earliest chronometrically dated Upper leys of south central Europe have yielded a Paleolithic-associated hominid in Europe large and significant series of Upper Pleisto- (Smith, 1976a). cene fossil hominids (e.g. Jelinek, 1969) as well This report presents a detailed comparative as extensive evidence of their cultural behavior description of a sample of fossil hominids re- (e.g. -

Chapter 4: Whai Can the Fossil Record Tell Lis About Human Origins?
anthropq what does it mean to be human? FOURTH EDITION Robert H. Lavenda St. Cloud State University Emily A. Schultz St. Cloud State University NewYork Oxford ^ot8OXFORD UNIVERSITY PRESS What can the fossil record tell us about human origi ns? Anthropology has made major contributions to our understanling of human biological and cultural evolution. This chapter tells the story of what we have learned from fossils, stone tools, and other cultural remains, from the appearance of our earliest known ancestors about 6 millioil years ago through the appearance of modern Homo sapiens about 200,000 years ago. CHAPTER OUTLINE What ls Macroevolution? The Culture of H. erectus What Do We Know about the What ls Hominin Evolution? H. erectusthe Hunter? Upper Paleolithic/Late Stone Who Were the First Hominins What Happened to H. erectus? Age (40,000?-1 2,000 Years Ago)? (6-3 mya)? How Did Homo sopiens Evolve? The Origin of Bipedalism What ls the Fossil Evidence for What Happened to the Changes in Hominin Dentition the Transition to Modern H. Neandertals? Who Were the Later sapiens? How Many Kinds of Upper Australopiths (3-1.5 mya)? Where Did Modern H. sapiens Paleolithic/Late Stone Age How Many Species of Come from? Cultures Were There? Australopith Were There? Who Were the Neandertals Where Did Modern H. sapiens How Can Anthropologists (1 30,000-35,000 Years Ago)? Migrate in Late Pleistocene Explain the Human Transition? What Do We Know about Times? What Do We Know about Early Middle Paleolithic/Middle Eastern Asia and Siberia Homo (2.4-1.5 mya)? -

Growth, Learning, Play and Attachment in Neanderthal Children
This is a repository copy of The Cradle of Thought : Growth, Learning, Play and Attachment in Neanderthal children. White Rose Research Online URL for this paper: https://eprints.whiterose.ac.uk/83027/ Version: Submitted Version Article: Spikins, Penny orcid.org/0000-0002-9174-5168, Hitchens, Gail, Rutherford, Holly et al. (1 more author) (2014) The Cradle of Thought : Growth, Learning, Play and Attachment in Neanderthal children. Oxford Journal of Archaeology. pp. 111-134. ISSN 0262-5253 https://doi.org/10.1111/ojoa.12030 Reuse Items deposited in White Rose Research Online are protected by copyright, with all rights reserved unless indicated otherwise. They may be downloaded and/or printed for private study, or other acts as permitted by national copyright laws. The publisher or other rights holders may allow further reproduction and re-use of the full text version. This is indicated by the licence information on the White Rose Research Online record for the item. Takedown If you consider content in White Rose Research Online to be in breach of UK law, please notify us by emailing [email protected] including the URL of the record and the reason for the withdrawal request. [email protected] https://eprints.whiterose.ac.uk/ THE CRADLE OF THOUGHT: GROWTH, LEARNING, PLAY AND ATTACHMENT IN NEANDERTHAL CHILDREN Penny Spikins, Gail Hitchens, Andy Needham and Holly Rutherford Department of Archaeology University of York King’s Manor York YO1 7EP SUMMARY Childhood is a core stage in development, essential in the acquisition of social, practical and cultural skills. However, this area receives limited attention in archaeological debate, especially in early prehistory. -

UNIT 4 HISTORY of HUMAN EVOLUTION* History of Human Evolution
UNIT 4 HISTORY OF HUMAN EVOLUTION* History of Human Evolution Contents 4.0 Introduction 4.1 Trends in Human Evolution: Understanding Pre-modern Humans 4.2 Hominization Process 4.2.1 Bipedalism 4.2.2 Opposable Thumb and Manual Dexterity 4.3 Summary 4.4 References 4.5 Answers to Check Your Progress Learning Objectives: After reading this unit you will be able to: analyze the major trends in human evolution; review characteristics which distinguish human from their primate ancestors; learn anatomical and cultural changes associated with the process of hominization; and comprehend the significance of these changes during evolution of human. 4.0 INTRODUCTION Humans first evolved in East Africa about 2.5 million years ago from an earlier genus of apes called Australopithecus, which means ‘Southern Ape’. About 2 million years ago, some of these archaic men and women left their homeland to journey through and settle vast areas of North Africa, Europe and Asia. Since survival in the snowy forests of northern Europe required different traits than those needed to stay alive in Indonesia’s steaming jungles, human populations evolved in different directions. The result was several distinct species, to each of which scientists have assigned a pompous Latin name. Humans in Europe and western Asia evolved into Homo neanderthalensis (‘Man from the Neander Valley’), popularly referred to simply as ‘Neandethals’. Neanderthals, bulkier and more muscular than us Sapiens, were well adapted to the cold climate of Ice Age western Eurasia. The more eastern regions of Asia were populated by Homo erects, ‘Upright Man’, who survived there for close to 2 million years, making it the most durable species ever. -

Mechanics of Bipedalism: an Exploration of Skeletal Morphology and Force Plate Anaylsis Erin Forse May 04, 2007 a Senior Thesis
MECHANICS OF BIPEDALISM: AN EXPLORATION OF SKELETAL MORPHOLOGY AND FORCE PLATE ANAYLSIS ERIN FORSE MAY 04, 2007 A SENIOR THESIS SUBMITTED IN PARTIAL FULFILLMENT OF THE REQUIREMENTS FOR THE DEGREE OF BACHELOR OF ARTS IN ARCHAEOLOGICAL STUDIES UNIVERSITY OF WISCONSIN- LA CROSSE Abstract There are several theories on how humans learned to walk, and while these all address the adaptations needed for walking, none adequately describes how our early ancestors developed the mechanism to walk. Our earliest recognizable relatives, the australopithecines, have several variations on a theme: walking upright. There are varied changes as australopithecines approach the genus Homo. These changes occurred in the spine, legs, pelvis, and feet, and changes are also in the cranium, arms and hands, but these are features that may have occurred simultaneously with bipedalism. Several analyses of Australopithecus afarensis, specifically specimen A.L. 288-1 ("Lucy"), have shown that the skeletal changes are intermediate between apes and humans. Force plate analyses are used to determine if the gait pattern of humans resembles that of apes, and if it is a likely development pattern. The results of both these analyses will give insight into how modern humans developed bipedalism. Introduction Bipedalism is classified as movement of the post-cranial body in a vertical position, with the lower limbs shifting as an inverted pendulum, progressing forward. Simply, it is upright walking. Several theories have addressed why bipedalism evolved in hominids, with some unlikely ideas taking hold throughout the history of the issue. Other theories are more likely, but all lack the same characteristic: answering how bipedalism developed. -

Early Evidence of Stone Tool Use in Bone Working Activities at Qesem Cave, Israel
www.nature.com/scientificreports OPEN Early evidence of stone tool use in bone working activities at Qesem Cave, Israel Received: 15 July 2016 Andrea Zupancich1, Stella Nunziante-Cesaro2, Ruth Blasco1,3, Jordi Rosell4,5, Accepted: 03 October 2016 Emanuela Cristiani6, Flavia Venditti7, Cristina Lemorini7, Ran Barkai1 & Avi Gopher1 Published: 25 November 2016 For a long while, the controversy surrounding several bone tools coming from pre-Upper Palaeolithic contexts favoured the view of Homo sapiens as the only species of the genus Homo capable of modifying animal bones into specialised tools. However, evidence such as South African Early Stone Age modified bones, European Lower Palaeolithic flaked bone tools, along with Middle and Late Pleistocene bone retouchers, led to a re-evaluation of the conception of Homo sapiens as the exclusive manufacturer of specialised bone tools. The evidence presented herein include use wear and bone residues identified on two flint scrapers as well as a sawing mark on a fallow deer tibia, not associated with butchering activities. Dated to more than 300 kya, the evidence here presented is among the earliest related to tool-assisted bone working intended for non-dietary purposes, and contributes to the debate over the recognition of bone working as a much older behaviour than previously thought. The results of this study come from the application of a combined methodological approach, comprising use wear analysis, residue analysis, and taphonomy. This approach allowed for the retrieval of both direct and indirect evidence of tool-assisted bone working, at the Lower Palaeolithic site of Qesem Cave (Israel). Homo sapiens’ supposedly exclusive manufacture of specialised tools made from modified animal bones, along with other aspects such as art and specialised hunting weapons, has led to the definition of a clear behavioural and cognitive boundary between H. -

Universidad Complutense De Madrid
UNIVERSIDAD COMPLUTENSE DE MADRID FACULTAD DE CIENCIAS BIOLÓGICAS Departamento de Zoología y Antropología Física TESIS DOCTORAL Las poblaciones del Holoceno inicial en la región cantábrica: cambios ambientales y microevolución humana MEMORIA PARA OPTAR AL GRADO DE DOCTOR PRESENTADA POR Labib Drak Hernández Directora María Dolores Garralda Benajes Madrid, 2016 © Labib Drak Hernández, 2016 UNIVERSIDAD COMPLUTENSE DE MADRID FACULTAD DE CIENCIAS BIOLÓGICAS DEPARTAMENTO DE ZOOLOGÍA Y ANTROPOLOGÍA FÍSICA TESIS DOCTORAL LAS POBLACIONES DEL HOLOCENO INICIAL EN LA REGIÓN CANTÁBRICA: CAMBIOS AMBIENTALES Y MICROEVOLUCIÓN HUMANA MEMORIA PARA OPTAR AL GRADO DE DOCTOR PRESENTADA POR LABIB DRAK HERNÁNDEZ BAJO LA DIRECCIÓN DE LA DOCTORA: MARÍA DOLORES GARRALDA BENAJES MADRID, 2015 UNIVERSIDAD COMPLUTENSE DE MADRID FACULTAD DE CIENCIAS BIOLÓGICAS DEPARTAMENTO DE ZOOLOGÍA Y ANTROPOLOGÍA FÍSICA TESIS DOCTORAL LAS POBLACIONES DEL HOLOCENO INICIAL EN LA REGIÓN CANTÁBRICA: CAMBIOS AMBIENTALES Y MICROEVOLUCIÓN HUMANA MEMORIA PARA OPTAR AL GRADO DE DOCTOR PRESENTADA POR LABIB DRAK HERNÁNDEZ BAJO LA DIRECCIÓN DE LA DOCTORA: MARÍA DOLORES GARRALDA BENAJES MADRID, 2015 MARÍA DOLORES GARRALDA BENAJES, PROFESORA TITULAR DEL DEPARTAMENTO DE ZOOLOGÍA Y ANTROPOLOGÍA FÍSICA DE LA FACULTAD DE CIENCIAS BIOLÓGICAS DE LA UNIVERSIDAD COMPLUTENSE DE MADRID, CERTIFICA: Que la presente memoria titulada “Las poblaciones del Holoceno inicial en la región cantábrica: cambios ambientales y microevolución humana” presentada por D. Labib Drak Hernández para optar al Título de Doctor en Biología, ha sido realizada en el Departamento de Zoología y Antropología Física de la Facultad de CC. Biológicas de la Universidad Complutense de Madrid bajo mi dirección. Y considerando que representa trabajo de Tesis, autorizo su presentación a la Junta de Facultad.