Platanus Wrightii)
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Arizona Fishing Regulations 3 Fishing License Fees Getting Started
2019 & 2020 Fishing Regulations for your boat for your boat See how much you could savegeico.com on boat | 1-800-865-4846insurance. | Local Offi ce geico.com | 1-800-865-4846 | Local Offi ce See how much you could save on boat insurance. Some discounts, coverages, payment plans and features are not available in all states or all GEICO companies. Boat and PWC coverages are underwritten by GEICO Marine Insurance Company. GEICO is a registered service mark of Government Employees Insurance Company, Washington, D.C. 20076; a Berkshire Hathaway Inc. subsidiary. TowBoatU.S. is the preferred towing service provider for GEICO Marine Insurance. The GEICO Gecko Image © 1999-2017. © 2017 GEICO AdPages2019.indd 2 12/4/2018 1:14:48 PM AdPages2019.indd 3 12/4/2018 1:17:19 PM Table of Contents Getting Started License Information and Fees ..........................................3 Douglas A. Ducey Governor Regulation Changes ...........................................................4 ARIZONA GAME AND FISH COMMISSION How to Use This Booklet ...................................................5 JAMES S. ZIELER, CHAIR — St. Johns ERIC S. SPARKS — Tucson General Statewide Fishing Regulations KURT R. DAVIS — Phoenix LELAND S. “BILL” BRAKE — Elgin Bag and Possession Limits ................................................6 JAMES R. AMMONS — Yuma Statewide Fishing Regulations ..........................................7 ARIZONA GAME AND FISH DEPARTMENT Common Violations ...........................................................8 5000 W. Carefree Highway Live Baitfish -

Water Resources of Bill Williams River Valley Near Alamo, Arizona
Water Resources of Bill Williams River Valley Near Alamo, Arizona GEOLOGICAL SURVEY WATER-SUPPLY PAPER 1360-D DEC 10 1956 Water Resources of Bill Williams River Valley Near Alamo, Arizona By H. N. WOLCOTT, H. E. SKIBITZKE, and L. C. HALPENNY CONTRIBUTIONS TO THE HYDROLOGY OF THE UNITED STATES GEOLOGICAL SURVEY WATER-SUPPLY PAPER 1360-D An investigation of the availability of water in the area of the Artillery Mountains manganese deposits UNITED STATES GOVERNMENT PRINTING OFFICE, WASHINGTON : 1956 UNITED STATES DEPARTMENT OF THE INTERIOR Fred A. Seaton, Secretary GEOLOGICAL SURVEY Thomas B. Nolan, Director For sale by the Superintendent of Documents, U. S. Government Printing Office Washington 25, D. C. - Price 45 cents (paper cover) CONTENTS Page Abstract....................................................................................................................................... 291 Introduction................................................................................................................................. 292 Purpose.................................................................................................................................... 292 Location................................................................................................................................... 292 Climatological data............................................................................................................... 292 History of development........................................................................................................ -

On the Pima County Multi-Species Conservation Plan, Arizona
United States Department of the Interior Fish and ,Vildlife Service Arizona Ecological Services Office 2321 West Royal Palm Road, Suite 103 Phoenix, Arizona 85021-4951 Telephone: (602) 242-0210 Fax: (602) 242-2513 In reply refer to: AESO/SE 22410-2006-F-0459 April 13, 2016 Memorandum To: Regional Director, Fish and Wildlife Service, Albuquerque, New Mexico (ARD-ES) (Attn: Michelle Shaughnessy) Chief, Arizona Branch, Re.. gul 7/to . D'vision, Army Corps of Engineers, Phoenix, Arizona From: Acting Field Supervisor~ Subject: Biological and Conference Opinion on the Pima County Multi-Species Conservation Plan, Arizona This biological and conference opinion (BCO) responds to the Fish and Wildlife Service (FWS) requirement for intra-Service consultation on the proposed issuance of a section lO(a)(l)(B) incidental take permit (TE-84356A-O) to Pima County and Pima County Regional Flood Control District (both herein referenced as Pima County), pursuant to section 7 of the Endangered Species Act of 1973 (U.S.C. 1531-1544), as amended (ESA), authorizing the incidental take of 44 species (4 plants, 7 mammals, 8 birds, 5 fishes, 2 amphibians, 6 reptiles, and 12 invertebrates). Along with the permit application, Pima County submitted a draft Pima County Multi-Species Conservation Plan (MSCP). On June 10, 2015, the U.S. Army Corps of Engineers (ACOE) requested programmatic section 7 consultation for actions under section 404 of the Clean Water Act (CW A), including two Regional General Permits and 16 Nationwide Permits, that are also covered activities in the MSCP. This is an action under section 7 of the ESA that is separate from the section 10 permit issuance to Pima Couny. -

Vascular Plant and Vertebrate Inventory of Chiricahua National Monument
In Cooperation with the University of Arizona, School of Natural Resources Vascular Plant and Vertebrate Inventory of Chiricahua National Monument Open-File Report 2008-1023 U.S. Department of the Interior U.S. Geological Survey National Park Service This page left intentionally blank. In cooperation with the University of Arizona, School of Natural Resources Vascular Plant and Vertebrate Inventory of Chiricahua National Monument By Brian F. Powell, Cecilia A. Schmidt, William L. Halvorson, and Pamela Anning Open-File Report 2008-1023 U.S. Geological Survey Southwest Biological Science Center Sonoran Desert Research Station University of Arizona U.S. Department of the Interior School of Natural Resources U.S. Geological Survey 125 Biological Sciences East National Park Service Tucson, Arizona 85721 U.S. Department of the Interior DIRK KEMPTHORNE, Secretary U.S. Geological Survey Mark Myers, Director U.S. Geological Survey, Reston, Virginia: 2008 For product and ordering information: World Wide Web: http://www.usgs.gov/pubprod Telephone: 1-888-ASK-USGS For more information on the USGS-the Federal source for science about the Earth, its natural and living resources, natural hazards, and the environment: World Wide Web:http://www.usgs.gov Telephone: 1-888-ASK-USGS Suggested Citation Powell, B.F., Schmidt, C.A., Halvorson, W.L., and Anning, Pamela, 2008, Vascular plant and vertebrate inventory of Chiricahua National Monument: U.S. Geological Survey Open-File Report 2008-1023, 104 p. [http://pubs.usgs.gov/of/2008/1023/]. Cover photo: Chiricahua National Monument. Photograph by National Park Service. Note: This report supersedes Schmidt et al. (2005). Any use of trade, product, or firm names is for descriptive purposes only and does not imply endorsement by the U.S. -

A Global Phylogeny of Leafmining Ectoedemia Moths (Lepidoptera: Nepticulidae): Host Plant Family Shifts and Allopatry As Drivers of Speciation
UvA-DARE (Digital Academic Repository) A global phylogeny of leafmining Ectoedemia moths (Lepidoptera: Nepticulidae): host plant family shifts and allopatry as drivers of speciation Doorenweerd, C.; van Nieukerken, E.J.; Menken, S.B.J. DOI 10.1371/journal.pone.0119586 Publication date 2015 Document Version Final published version Published in PLoS ONE License CC BY Link to publication Citation for published version (APA): Doorenweerd, C., van Nieukerken, E. J., & Menken, S. B. J. (2015). A global phylogeny of leafmining Ectoedemia moths (Lepidoptera: Nepticulidae): host plant family shifts and allopatry as drivers of speciation. PLoS ONE, 10(3), [e0119586]. https://doi.org/10.1371/journal.pone.0119586 General rights It is not permitted to download or to forward/distribute the text or part of it without the consent of the author(s) and/or copyright holder(s), other than for strictly personal, individual use, unless the work is under an open content license (like Creative Commons). Disclaimer/Complaints regulations If you believe that digital publication of certain material infringes any of your rights or (privacy) interests, please let the Library know, stating your reasons. In case of a legitimate complaint, the Library will make the material inaccessible and/or remove it from the website. Please Ask the Library: https://uba.uva.nl/en/contact, or a letter to: Library of the University of Amsterdam, Secretariat, Singel 425, 1012 WP Amsterdam, The Netherlands. You will be contacted as soon as possible. UvA-DARE is a service provided by the library of the University of Amsterdam (https://dare.uva.nl) Download date:28 Sep 2021 RESEARCH ARTICLE A Global Phylogeny of Leafmining Ectoedemia Moths (Lepidoptera: Nepticulidae): Exploring Host Plant Family Shifts and Allopatry as Drivers of Speciation Camiel Doorenweerd1,2*, Erik J. -
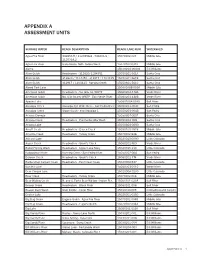
Appendix a Assessment Units
APPENDIX A ASSESSMENT UNITS SURFACE WATER REACH DESCRIPTION REACH/LAKE NUM WATERSHED Agua Fria River 341853.9 / 1120358.6 - 341804.8 / 15070102-023 Middle Gila 1120319.2 Agua Fria River State Route 169 - Yarber Wash 15070102-031B Middle Gila Alamo 15030204-0040A Bill Williams Alum Gulch Headwaters - 312820/1104351 15050301-561A Santa Cruz Alum Gulch 312820 / 1104351 - 312917 / 1104425 15050301-561B Santa Cruz Alum Gulch 312917 / 1104425 - Sonoita Creek 15050301-561C Santa Cruz Alvord Park Lake 15060106B-0050 Middle Gila American Gulch Headwaters - No. Gila Co. WWTP 15060203-448A Verde River American Gulch No. Gila County WWTP - East Verde River 15060203-448B Verde River Apache Lake 15060106A-0070 Salt River Aravaipa Creek Aravaipa Cyn Wilderness - San Pedro River 15050203-004C San Pedro Aravaipa Creek Stowe Gulch - end Aravaipa C 15050203-004B San Pedro Arivaca Cienega 15050304-0001 Santa Cruz Arivaca Creek Headwaters - Puertocito/Alta Wash 15050304-008 Santa Cruz Arivaca Lake 15050304-0080 Santa Cruz Arnett Creek Headwaters - Queen Creek 15050100-1818 Middle Gila Arrastra Creek Headwaters - Turkey Creek 15070102-848 Middle Gila Ashurst Lake 15020015-0090 Little Colorado Aspen Creek Headwaters - Granite Creek 15060202-769 Verde River Babbit Spring Wash Headwaters - Upper Lake Mary 15020015-210 Little Colorado Babocomari River Banning Creek - San Pedro River 15050202-004 San Pedro Bannon Creek Headwaters - Granite Creek 15060202-774 Verde River Barbershop Canyon Creek Headwaters - East Clear Creek 15020008-537 Little Colorado Bartlett Lake 15060203-0110 Verde River Bear Canyon Lake 15020008-0130 Little Colorado Bear Creek Headwaters - Turkey Creek 15070102-046 Middle Gila Bear Wallow Creek N. and S. Forks Bear Wallow - Indian Res. -

Histoire Naturelle Du Genre De Fourmis Néotropical Thaumatomyrmex Mayr 1887 (Arthropoda, Insecta, Hymenoptera, Formicidae, Ponerinae, Thaumatomyrmecini)
UNIVERSITE PARIS XIII ________________________________________ École Doctorale de Galilée THÈSE Présentée pour obtenir le grade de DOCTEUR DE L'UNIVERSITÉ PARIS XIII Discipline : Éthologie Par Benoit Jean Bernard JAHYNY VOLUME 1 / 2 Histoire Naturelle du genre de fourmis néotropical Thaumatomyrmex Mayr 1887 (Arthropoda, Insecta, Hymenoptera, Formicidae, Ponerinae, Thaumatomyrmecini) présentée et soutenue le 28 janvier 2010 devant le jury composé de: P. JAISSON Professeur Président (Université Paris XIII, Villetaneuse) A. LENOIR Professeur émérite Rapporteur (Université François Rabelais, Tours) C. PEETERS Directeur de Recherche CNRS Rapporteur (Université Paris VI) S. LACAU Professeur Examinateur (UESB, Itapetinga, BA, Brésil) J. ORIVEL Chercheur CNRS Examinateur (Université Paul Sabatier, Toulouse) D. FRESNEAU Professeur Examinateur (Université Paris XIII, Villetaneuse) (Codirecteur) J.H.C. DELABIE Professeur Examinateur (UESC - CEPEC/CEPLAC, Itabuna, BA, Brésil) (Codirecteur) Laboratoire d’Ethologie Expérimentale et Comparée (EA 4443) et Laboratório de Mirmecologia, Centro de Pesquisa do Cacau (CEPEC/CEPLAC) À mes parents, ils comprendront, e à Raquel, ela entenderá 2 Remerciements Ce furent sept ans de dur labeur, de travail dans l’ombre, de longues lectures et réflexions, d’essais/erreurs, de frustrations, de désillusions, de malchance, de dispersion (un peu) ; les motifs de joies et de satisfactions furent peu nombreux, mais toujours très intenses et je n’ai jamais abandonné car si « impossible n’est pas français, « sou brasileiro abaianizado de adopção e por isso desisto nunca ! ». Tout cela ne s’est pas fait sans l’apport des amitiés anciennes ou nouvelles que j’ai pu nouer, ni sans l’aide indéfectible de ma famille. J’aimerais remercier simplement : les membres du jury pour avoir accepté de corriger ce manuscrit en deux volumes. -

Ecological Site R041XC316AZ Sandy Wash 12-16" P.Z
Natural Resources Conservation Service Ecological site R041XC316AZ Sandy Wash 12-16" p.z. Accessed: 09/23/2021 General information Provisional. A provisional ecological site description has undergone quality control and quality assurance review. It contains a working state and transition model and enough information to identify the ecological site. Figure 1. Mapped extent Areas shown in blue indicate the maximum mapped extent of this ecological site. Other ecological sites likely occur within the highlighted areas. It is also possible for this ecological site to occur outside of highlighted areas if detailed soil survey has not been completed or recently updated. MLRA notes Major Land Resource Area (MLRA): 041X–Southeastern Arizona Basin and Range AZ 41.3 – Chihuahuan – Sonoran Semidesert Grasslands Elevations range from 3200 to 5000 feet and precipitation ranges from 12 to 16 inches per year. Vegetation includes mesquite, catclaw acacia, netleaf hackberry, palo verde, false mesquite, range ratany, fourwing saltbush, tarbush, littleleaf sumac, sideoats grama, black grama, plains lovegrass, cane beardgrass, tobosa, vine mesquite, threeawns, Arizona cottontop and bush muhly. The soil temperature regime is thermic and the soil moisture regime is ustic aridic. This unit occurs within the Basin and Range Physiographic Province and is characterized by numerous mountain ranges that rise abruptly from broad, plain-like valleys and basins. Igneous and metamorphic rock classes dominate the mountain ranges and sediments filling the basins represent combinations of fluvial, lacustrine, colluvial and alluvial deposits. Associated sites R041XC306AZ Shallow Hills 12-16" p.z. R041XC308AZ Limy Slopes 12-16" p.z. R041XC309AZ Limy Upland 12-16" p.z. R041XC313AZ Loamy Upland 12"-16" p.z. -

A Global Phylogeny of Leafmining Ectoedemia Moths (Lepidoptera: Nepticulidae): Exploring Host Plant Family Shifts and Allopatry As Drivers of Speciation
RESEARCH ARTICLE A Global Phylogeny of Leafmining Ectoedemia Moths (Lepidoptera: Nepticulidae): Exploring Host Plant Family Shifts and Allopatry as Drivers of Speciation Camiel Doorenweerd1,2*, Erik J. van Nieukerken1, Steph B. J. Menken2 1 Department of Terrestrial Zoology, Naturalis Biodiversity Center, Leiden, The Netherlands, 2 Institute for Biodiversity and Ecosystem Dynamics, University of Amsterdam, Amsterdam, The Netherlands a11111 * [email protected] Abstract OPEN ACCESS Background Citation: Doorenweerd C, van Nieukerken EJ, Host association patterns in Ectoedemia (Lepidoptera: Nepticulidae) are also encountered Menken SBJ (2015) A Global Phylogeny of in other insect groups with intimate plant relationships, including a high degree of monopha- Ectoedemia Leafmining Moths (Lepidoptera: gy, a preference for ecologically dominant plant families (e.g. Fagaceae, Rosaceae, Salica- Nepticulidae): Exploring Host Plant Family Shifts and Allopatry as Drivers of Speciation. PLoS ONE 10(3): ceae, and Betulaceae) and a tendency for related insect species to feed on related host e0119586. doi:10.1371/journal.pone.0119586 plant species. The evolutionary processes underlying these patterns are only partly under- Academic Editor: William J. Etges, University of stood, we therefore assessed the role of allopatry and host plant family shifts in speciation Arkansas, UNITED STATES within Ectoedemia. Received: July 15, 2014 Methodology Accepted: January 14, 2015 Six nuclear and mitochondrial DNA markers with a total aligned length of 3692 base pairs Published: March 18, 2015 were used to infer phylogenetic relationships among 92 species belonging to the subgenus Copyright: © 2015 Doorenweerd et al. This is an Ectoedemia of the genus Ectoedemia, representing a thorough taxon sampling with a global open access article distributed under the terms of the Creative Commons Attribution License, which permits coverage. -

The Colorado River: Lifeline Of
4 The Colorado River: lifeline of the American Southwest Clarence A. Carlson Department of Fishery and Wildlife Biology, Colorado State University, Fort Collins, CO, USA 80523 Robert T. Muth Larval Fish Laboratory, Colorado State University, Fort Collins, CO, USA 80523 1 Carlson, C. A., and R. T. Muth. 1986. The Colorado River: lifeline of the American Southwest. Can. J. Fish. Aguat. Sci. In less than a century, the wild Colorado River has been drastically and irreversibly transformed into a tamed, man-made system of regulated segments dominated by non-native organisms. The pristine Colorado was characterized by widely fluctuating flows and physico-chemical extremes and harbored unique assemblages of indigenous flora and fauna. Closure of Hoover Dam in 1935 marked the end of the free-flowing river. The Colorado River System has since become one of the most altered and intensively controlled in the United States. Many main-stem and tributary dams, water diversions, and channelized river sections now exist in the basin. Despite having one of - the most arid drainages in the world, the present-day Colorado River supplies more water for consumptive use than any river in the United States. Physical modification of streams and introduction of non-native species have adversely impacted the Colorado's native biota. This paper treats the Colorado River holistically as an ecosystem and summarizes current knowledge on its ecology and management. "In a little over two generations, the wild Colorado has been harnessed by a series of dams strung like beads on a thread from the Gulf of California to the mountains of Wyoming. -

Analysis of Sediment Dynamics in the Bill Williams River, Arizona
Analysis of sediment dynamics in the Bill Williams River, Arizona Andrew C. Wilcox and Franklin Dekker Department of Geosciences Center for Riverine Science and Stream Renaturalization University of Montana Missoula, MT 59812-1296 Paul Gremillion (Northern Arizona University) and David Walker (University of Arizona) Collaborators: Patrick Shafroth (USGS), Cliff Riebe (University of Wyoming), Kyle House (USGS), John Stella (State University of New York) Report prepared for U.S. Fish and Wildlife Service, via Rocky Mountains Cooperative Ecosystem Studies Unit (RM-CESU) February 6, 2013 DRAFT FINAL REPORT Table of Contents Executive summary ......................................................................................................................... 2 Introduction .................................................................................................................................... 4 Catchment Erosion Rates and Sediment Mixing in a Dammed Dryland River ............................. 11 Dryland River Grain-Size Variation due to Damming, Tributary Confluences, and Valley Confinement ................................................................................................................................. 28 Analysis of Sediment Dynamics in the Bill Williams River, Arizona: Hydroacoustic Surveys and Sediment Coring ............................................................................................................................ 46 Coupled Hydrogeomorphic and Woody-Seedling Responses to Controlled Flood -

Salinity of Surface Water in the Lower Colorado River Salton Sea Area
Salinity of Surface Water in The Lower Colorado River Salton Sea Area GEOLOGICAL SURVEY PROFESSIONAL PAPER 486-E Salinity of Surface Water in The Lower Colorado River- Salton Sea Area By BURDGE IRELAN WATER RESOURCES OF LOWER COLORADO RIVER SALTON SEA AREA GEOLOGICAL SURVEY PROFESSIONAL PAPER 486-E UNITED STATES GOVERNMENT PRINTING OFFICE, WASHINGTON : 1971 UNITED STATES DEPARTMENT OF THE INTERIOR ROGERS C. B. MORTON, Secretary GEOLOGICAL SURVEY William T. Pecora, Director Library of Congress catalog-card No. 72 610761 For sale by the Superintendent of Documents, U.S. Government Printing Office Washington, D.C. 20402 Price 50 cents (paper cover) CONTENTS Page Page Abstract . _.._.-_. ._...._ ..._ _-...._ ...._. ._.._... El Ionic budget of the Colorado River from Lees Ferry to Introduction .._____. ..... .._..__-. - ._...-._..__..._ _.-_ ._... 2 Imperial Dam, 1961-65 Continued General chemical characteristics of Colorado River Tapeats Creek .._________________.____.___-._____. _ E26 water from Lees Ferry to Imperial Dam ____________ 2 Havasu Creek __._____________-...- _ __ -26 Lees Ferry .._._..__.___.______.__________ 4 Virgin River ..__ .-.._..-_ --....-. ._. 26 Grand Canyon ................._____________________..............._... 6 Unmeasured inflow between Grand Canyon and Hoover Dam ..........._._..- -_-._-._................-._._._._... 8 Hoover Dam .__-.....-_ .... .-_ . _. 26 Lake Havasu - -_......_....-..-........ .........._............._.... 11 Chemical changes in Lake Mead ............-... .-.....-..... 26 Imperial Dam .--. ........_. ...___.-_.___ _.__.__.._-_._.___ _ 12 Bill Williams River ......._.._......__.._....._ _......_._- 27 Mineral burden of the lower Colorado River, 1926-65 .