Rogan Et Al 2017 Illegal Bushmeat Hunters Compete with Predators And
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Okavango River Basin Groundwater Overview
Okavango River Basin Groundwater Overview Specialist Report prepared by Interconsult Namibia for : PERMANENT OKAVANGO RIVER BASIN COMMISSION Angola Botswana Namibia Ministério da Energia e Águas Ministry of Mineral Resources and Water Affairs Ministry of Agriculture, Water and Rural Development GABHIC Department of Water Affairs Department of Water Affairs Cx. P. 6695 Private Bag 0029 Private Bag 13193 LUANDA GABORONE WINDHOEK Tel: +244 2 393 681 Tel: +267 360 7100 Tel: +264 61 296 9111 Fax: +244 2 393 687 Fax: +267 303508 Fax: +264 61 232 861 PERMANENT OKAVANGO RIVER BASIN COMMISSION (OKACOM) OKAVANGO RIVER BASIN PREPARATORY ASSESSMENT: GROUNDWATER OVERVIEW Report prepared by: Interconsult Namibia (Pty) Ltd P. O. Box 20690 Windhoek With input from Wellfield Consulting Services, and E. Bereslawski March 1999 TABLE OF CONTENTS 1 INTRODUCTION ............................................................................................................... 1 1.1. BACKGROUND.............................................................................................................. 1 1.2. TERMS OF REFERENCE ................................................................................................ 1 2 OVERVIEW OF THE GEOHYDROLOGY OF THE BASIN .............................................. 2 2.1 BASIN GEOLOGY.............................................................................................................. 2 2.2 GEOHYDROLOGY OVERVIEW ........................................................................................... -

A Baseline Analysis of Poaching in Chiquibul National Park
A BASELINE ANALYSIS OF POACHING IN CHIQUIBUL NATIONAL PARK BY KATHERINE GROFF A THESIS Submitted to Michigan State University in partial fulfillment of the requirements for the degree of Master of Science Fisheries and Wildlife ABSTRACT A BASELINE ANALYSIS OF POACHING IN CHIQUIBUL NATIONAL PARK BY KATHERINE GROFF Illegal hunting of wildlife, also known as poaching, in Chiquibul National Park (CNP) may be contributing to decreasing wildlife populations. Management strategies are limited due to a lack of information on the extent of poaching and motivations of poachers. The objectives of this research are to assess the extent of poaching, to address Guatemalan border community residents’ understanding of wildlife in CNP and what animal species are targeted by hunters, and to investigate factors affecting poaching in CNP and Guatemalan border residents’ perceptions about poaching in CNP. These objectives were addressed by (1) synthesizing information found in CNP Ranger field notebooks and reports, (2) interviewing Guatemalan border community residents, and (3) interviewing authorities in Guatemala and Belize. Guatemalan border community residents perceived a lower level of poaching in the CNP than CNP rangers, but both thought that poaching has decreased over the years. CNP rangers considered poaching to be a danger to wildlife in the area, and they thought that wildlife numbers have suffered due to poaching. This research demonstrated little support for the hypothesis that legal regulations and the enforcement of regulations are factors that contribute to whether respondents view other community members as contributing to poaching. However, the subsistence needs of hunters and their families is an important factor affecting residents’ decision to poach. -

Illegal and Unsustainable Hunting of Wildlife for Bushmeat in Sub-Saharan Africa
About the Wilderness Problem-Specific Guide Series These guides summarize knowledge about how wildlife authorities can reduce the harm caused by specific wildlife crime problems. They are guides to preventing and improving the overall response to incidents, not to investigating offenses or handling specific incidents; neither do they cover technical details about how to implement specific responses. Who is this bushmeat guide for? This guide is aimed at wildlife officers and non-governmental conservation practitioners who have identified the illegal and unsustainable hunting of wildlife for bushmeat, as an important threat in a specific site or landscape. These include: ñ Protected Area Managers and their deputies ñ Conservation NGO Project Leads ñ Wildlife officers and NGO conservation practitioners of whatever rank or assignment, who have been tasked to address the problem These guides will be most useful to problem solvers who: Understand basic problem-oriented policing principles and methods. The guides are designed to help conservation practitioners decide how best to analyze Scanning Analysis Collect and analyze and address a problem they have already Identify and prioritize information to determine problems. Choose one what drives and facilitates identified. The guides are structured in specific problem. the same way as the SARA process the problem. (right). This covers how to define your problem (Scan); questions you will need to answer to guide you to an effective intervention (Analysis); types of interventions you could use (Response); and ways to check if your intervention worked (Assessment). Response Assessment Implement response that reduces drivers and For a primer on Problem-Oriented Determine the impact of your facilitators of the problem. -

Kōji-Poached Chicken
麹漬け鶏肉の酒蒸し Kōji-zuké Toriniku no Saka Mushi KŌJI-POACHED CHICKEN The Japanese cooking technique known as SAKA MUSHI literally “saké steamed” is the equivalent of poaching. The technique is most often used to cook fish, shellfish or chicken; it keeps the food moist as it gently cooks in a limited amount of saké and broth. Here chicken breasts, or thighs, are briefly, lightly massaged in komé kōji (malted rice) first; this softens otherwise tough muscle tissue and sinews. And, it makes the chicken less “gamey” tasting, too. Making this dsh can be worked into even the busiest of schedules. Massage and marinate the chicken in the refrigerator in the morning and finish by poaching in a shallow pan in the evening. Once the chicken has been prepared and cooked it will keep in the refrigerator for 2-3 days. The poached meat can be sliced and served with various sauces or condiments. Or shred and add to salads, or use to top noodles. It makes great sandwich meat, too. I urge you to leave the skin on while marinating and poaching the chicken to keep the meat moist and flavorful. You can remove it later if you prefer not to eat it. Boneless breast meat with skin (upper right) Boneless thigh meat skin side down (bottom right) © Copyright 2021 All rights reserved by Elizabeth Andoh 1 Makes enough chicken to top 4 noodle dishes or salads. 1 large boneless breast OR thigh with skin attached (about 12 oz/340 grams) 1 teaspoon komé kōji (malted rice) 1 strip kombu (kelp) soaked in 1 and 1/2 cups water for 20 minutes or longer 2 tablespoons saké With the tines of a fork, poke through the skin of the chicken breast – this will permit the flavor-enhancing and tenderizing properties of the kōji to work more efficiently. -

CHAPTER-2 Charcutierie Introduction: Charcuterie (From Either the French Chair Cuite = Cooked Meat, Or the French Cuiseur De
CHAPTER-2 Charcutierie Introduction: Charcuterie (from either the French chair cuite = cooked meat, or the French cuiseur de chair = cook of meat) is the branch of cooking devoted to prepared meat products such as sausage primarily from pork. The practice goes back to ancient times and can involve the chemical preservation of meats; it is also a means of using up various meat scraps. Hams, for instance, whether smoked, air-cured, salted, or treated by chemical means, are examples of charcuterie. The French word for a person who prepares charcuterie is charcutier , and that is generally translated into English as "pork butcher." This has led to the mistaken belief that charcuterie can only involve pork. The word refers to the products, particularly (but not limited to) pork specialties such as pâtés, roulades, galantines, crépinettes, etc., which are made and sold in a delicatessen-style shop, also called a charcuterie." SAUSAGE A simple definition of sausage would be ‘the coarse or finely comminuted (Comminuted means diced, ground, chopped, emulsified or otherwise reduced to minute particles by mechanical means) meat product prepared from one or more kind of meat or meat by-products, containing various amounts of water, usually seasoned and frequently cured .’ A sausage is a food usually made from ground meat , often pork , beef or veal , along with salt, spices and other flavouring and preserving agents filed into a casing traditionally made from intestine , but sometimes synthetic. Sausage making is a traditional food preservation technique. Sausages may be preserved by curing , drying (often in association with fermentation or culturing, which can contribute to preservation), smoking or freezing. -

Okavango Delta - 2020 Conservation Outlook Assessment
IUCN World Heritage Outlook: https://worldheritageoutlook.iucn.org/ Okavango Delta - 2020 Conservation Outlook Assessment Okavango Delta 2020 Conservation Outlook Assessment SITE INFORMATION Country: Botswana Inscribed in: 2014 Criteria: (vii) (ix) (x) This delta in north-west Botswana comprises permanent marshlands and seasonally flooded plains. It is one of the very few major interior delta systems that do not flow into a sea or ocean, with a wetland system that is almost intact. One of the unique characteristics of the site is that the annual flooding from the River Okavango occurs during the dry season, with the result that the native plants and animals have synchronized their biological cycles with these seasonal rains and floods. It is an exceptional example of the interaction between climatic, hydrological and biological processes. The Okavango Delta is home to some of the world’s most endangered species of large mammal, such as the cheetah, white rhinoceros, black rhinoceros, African wild dog and lion. © UNESCO SUMMARY 2020 Conservation Outlook Finalised on 01 Dec 2020 GOOD WITH SOME CONCERNS The nature of the Okavango Delta – a vast inaccessible wetland on the fringes of a sparsely populated desert – gives it a high degree of natural protection. The values of the site remain in good condition overall, largely due to the sites large size and inaccessibility, allowing the site to maintain its largely unaltered, pristine condition through low human impact. Threats to the site are generally of low concern but for a few issues which remain challenging to address. Whilst tourism has some negative impacts, there are very few roads and the industry is built around a high-cost low-volume business model with small lodge facilities accessed by private charter aircraft. -

Mega Kalahari Geology: Challenges of Kimberlite Exploration in This Medium
MEGA KALAHARI GEOLOGY: CHALLENGES OF KIMBERLITE EXPLORATION IN THIS MEDIUM Clint Williams1, Brett van Coller1, Tom Nowicki2 and John Gurney1 1 Mineral Services South Africa Pty Ltd, Cape Town.; 2 Mineral Services Canada Inc, Vancouver. obstacles and challenges to the kimberlite explorationist INTRODUCTION attempting to locate bedrock-hosted diamondiferous kimberlite bodies. The paper presents the complexities of this cover sequence, with a view to formulating In southern Africa the name Kalahari refers to a improved kimberlite exploration strategies in this structural basin, a group of terrestrial continental environment. sediments and an ill-defined desert, all of which are intimately linked yet also possess distinct characteristics. The surface units of the Kalahari Group CHARACTERISTICS cover represent the worlds largest body of sand covering over 2.5 million km² of central and southern The Mega Kalahari sediments represent an ancient Africa from the Orange River at 29°S to 1°N in the depositional environment with a complex history in western Congo and southern Gabon (Thomas, which the stratigraphy and age of the deposits are not 1988),(Figure 1). Thomas and Shaw (1990) grouped all particularly well constrained or understood. Low fossil these sediments together under the term Mega Kalahari. content, limited exposure, poor differentiation of the This term has been adopted for the review. dominant surficial Kalahari Sand and a limited comprehension of an extensive duricrust suite has delayed the understanding of the sedimentological and environmental history of the basin. Collectively the sediments are referred to as the Kalahari Group (SACS, 1980). SACS (1980) distinguished 6 major lithological types in the Kalahari Group: conglomerate and gravel, marl, sandstone, alluvium and lacustrine deposits, Kalahari Sand, and duricrusts (mainly calcrete and silcrete). -

Dynamic Evolution of the Zambezi-Limpopo Watershed, Central Zimbabwe
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by Online Research @ Cardiff Moore, Blenkinsop and Cotterill Zim watershed_20120408_AEM DYNAMIC EVOLUTION OF THE ZAMBEZI-LIMPOPO WATERSHED, CENTRAL ZIMBABWE Andy Moore1,2, Tom Blenkinsop3 and Fenton (Woody) Cotterill4 1African Queen Mines Ltd., Box 66 Maun, Botswana. 2Dept of Geology, Rhodes University, Grahamstown 6140, South Africa. Email: [email protected] 3School of Earth and Environmental Sciences, James Cook University, Townsville, QLD4811, Australia. Email: [email protected] 4AEON – African Earth Observatory Network, Geoecodynamics Research Hub, University of Stellenbosch, Private Bag X1, Matieland 7602, South Africa. Email: [email protected] ABSTRACT Prospecting carried out to the south of the Zambezi-Limpopo drainage divide in the vicinity of Bulawayo, Zimbabwe, led to the recovery of a suite of ilmenites with a chemical “fingerprint” that can be closely matched with the population found in the early Palaeozoic Colossus kimberlite, which is located to the north of the modern watershed. The ilmenite geochemistry eliminates other Zimbabwe Kimberlites as potential sources of these pathfinder minerals. Geophysical modelling has been used to ascribe the elevation of southern Africa to dynamic topography sustained by a mantle plume; however, the evolution of the modern divide between the Zambezi and Limpopo drainage basins is not readily explained in terms of this model. Rather, it can be interpreted to represent a late Palaeogene continental flexure, which formed in response to crustal shortening, linked to intra-plate transmission of stresses associated with an episode of spreading reorganization at the ocean ridges surrounding southern Africa. -
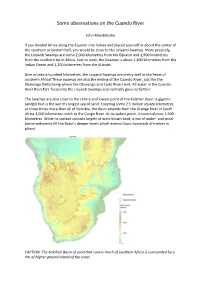
Some Observations on the Cuando River
Some observations on the Cuando River John Mendelsohn If you divided Africa along the Equator into halves and placed yourself at about the center of the southern or bottom half, you would be close to the Linyanti Swamps. More precisely, the Linyanti Swamps are some 2,000 kilometres from the Equator and 1,900 kilometres from the southern tip of Africa. East to west, the Swamps is about 1,400 kilometres from the Indian Ocean and 1,200 kilometres from the Atlantic. Give or take a hundred kilometres, the Linyanti Swamps are pretty well at the heart of southern Africa! These swamps are also the ending of the Cuando River, just like the Okavango Delta being where the Okavango and Cuito Rivers end. All water in the Cuando River therefore feeds into the Linyanti Swamps and normally goes no further. The Swamps are also close to the centre and lowest point of the Kalahari Basin: a gigantic sandpit that is the world’s largest sea of sand. Covering some 2.5 million square kilometres, or three times more than all of Namibia, the Basin extends from the Orange River in South Africa 3,000 kilometres north to the Congo River. At its widest point, it covers almost 1,500 kilometres. While its surface consists largely of wind‐blown sand, a mix of water‐ and wind‐ borne sediments fill the Basin’s deeper levels which extend down hundreds of metres in places. CAPTION: The Kalahari Basin of sand that covers much of southern Africa is surrounded by a rim of higher ground inland of the coast. -

COOKERY PROCESSES (COOKING METHODS) a Lot of Cooking
COOKERY PROCESSES (COOKING METHODS) A lot of cooking methods are used in catering and hotel industry. Each is specific and has its advantages and disadvantages. The cookery processes or cooking methods are: a) Boiling b) Poaching c) Stewing d) Braising e) Steaming f) Baking g) Roasting h) Pot roasting i) Grilling j) Shallow Frying k) Deep Frying l) Microwaving 1. Boiling www.astro.su.se/.../small_500/Boiling_water.jpg 1.1 Definition Boiling is cooking prepared foods in a liquid (water, bouillon, stock, milk) at boiling point. 1.2 Methods Food is boiled in two ways: a) food is placed into boiling liquid, reboiled, then the heat is reduced, so that the liquid boils gently – simmering; b) food is covered with cold liquid, brought to the boil, then the heat is reduced, so that the food simmers. 1.3 Advantages a) older, tougher joints of meat can be made palatable and digestible b) appropriate for large-scale cookery - 2 - c) economic on fuel d) nutritious, well flavoured stock is produced e) labor saving, requires little attention f) safe and simple g) maximum colour and nutritive value are retained with green vegetables – but the boiling time must be kept to the minimum 1.4 Disadvantages a) foods can look unattractive b) it can be slow c) loss of soluble vitamins in the water 1.5 Examples of foods which might be cooked by boiling - stocks (beef, mutton, chicken, fish) - sauces (brown, white, curry) - glazes (fish, meat) - soup (tomato, lentil) - farinaceous (pasta) - fish (cod, salmon) - meat (beef, leg of mutton) - vegetables (carrots, cabbage, potatoes). -

Sport Hunting in the Southern African Development Community (Sadc) Region
SPORT HUNTING IN THE SOUTHERN AFRICAN DEVELOPMENT COMMUNITY (SADC) REGION: An overview Rob Barnett Claire Patterson TRAFFIC East/Southern Africa Published by TRAFFIC East/Southern Africa, Johannesburg, South Africa. © 2006 TRAFFIC East/Southern Africa All rights reserved. All material appearing in this publication is copyrighted and may be reproduced with permission. Any reproduction in full or in part of this publication must credit TRAFFIC East/Southern Africa as the copyright owner. The views of the authors expressed in this publication do not necessarily reflect those of the TRAFFIC network, WWF or IUCN. The designations of geographical entities in this publication, and the presentation of the material, do not imply the expression of any opinion whatsoever on the part of TRAFFIC or its supporting organizations concerning the legal status of any country, territory, or area, or of its authorities, or concerning the delimitation of its frontiers or boundaries. The TRAFFIC symbol copyright and Registered Trademark ownership is held by WWF. TRAFFIC is a joint programme of WWF and IUCN. Suggested citation: Barnett, R. and Patterson, C. (2005). Sport Hunting in the Southern African Development Community ( SADC) Region: An overview. TRAFFIC East/Southern Africa. Johannesburg, South Africa ISBN: 0-9802542-0-5 Front cover photograph: Giraffe Giraffa camelopardalis Photograph credit: Megan Diamond Pursuant to Grant No. 690-0283-A-11-5950-00 Regional Networking and Capacity Building Initiative for southern Africa IUCN Regional Office for southern Africa “This publication was made possible through support provided by US Agency for International Development, REGIONAL CENTRE FOR SOUTHERN AFRICA under the terms of Grant No. -

Stocks, Broths, Stews & Soups
Stocks, Broths, Stews & Soups Greek Island Spice products are a Allow to simmer a few minutes or geat tool for creating flavorfl add to stock during simmer stage. soups and broths. Simply add Adjust to taste for a rich and aromatic beteen 1/4 to 1/2 ounce of base. Here are some ideas to keep your product per 8 oz. porion, or ketle a non stop culinar 4-8 oz. per gallon of adventre. prepared stock. Greek Island Spice Product Name Suggested Applications Ideas and Tips Add at the beginning of the cooking process, then Adobo Sauce Great starter for any Southwest soup, stew or broth. finish with additional to adjust heat level. Great seasoning base for a spicy broth for rice, Add an ounce of Jerk per gallon of liquid and a bit Caribbean Jerk Marinade grains, vegetables and stews of coconut for a perfect Jamaican Coconut Rice Classic South African Curry Stew, perfect for lamb, Durban Curry Use as marinade and to season stew as it cooks add a few apricots at the end for sweetness Use as marinade to braise meat for stew or add to Make an African themed chili with the addition of Ethiopian Berbere Marinade chili to add interesting twist root vegetables, beans and corn Moorish Spice Paste Great seasoning base for a Moorish style stew Red Snapper in Rich Moorish Spiced Broth Moroccan Chermoula Great seasoning base for a Moroccan style fish stew Moroccan Style Shrimp and Grouper Stew Add to broth or stock to impart traditional North Moroccan Spiced Heirloom rice and lentil pilaf Moroccan Steak Marinade African flavors with apricots and fresh mint