Voacanga Thou
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Studies on the Pharmacology of Conopharyngine, an Indole Alkaloid of the Voacanga Series
Br. J. Pharmac. Chemother. (1967), 30, 173-185. STUDIES ON THE PHARMACOLOGY OF CONOPHARYNGINE, AN INDOLE ALKALOID OF THE VOACANGA SERIES BY P. R. CARROLL AND G. A. STARMER From the Department of Pharmacology, University of Sydney, New South Wales, Australia (Received January 17, 1967) Conopharyngine, the major alkaloid present in the leaves of Tabernaemontana (Conopharyngia) pachysiphon var. cumminsi (Stapf) H. Huber was isolated and identified by Thomas & Starmer (1963). The same alkaloid has also been found in the stem bark of a Nigerian variety of the same species by Patel & Poisson (1966) and in the stem bark of Conopharyngia durissima by Renner, Prins & Stoll (1959). Conopharyn- gine is an indole alkaloid of the voacanga type, being 18-carbomethoxy-12,13- dimethoxyibogamine (Fig. 1) and is thus closely related to voacangine and coronaridine. Me.0OC Fig. 1. Conopharyngine (18-carbomethoxy-12,13-dimethoxyibogamine). Some confusion exists in that an alkaloid with an entirely different structure, but also named conopharyngine, was isolated from a cultivated variety of Conopharyngia pachysiphon by Dickel, Lucas & Macphillamy (1959). This compound was shown to be the 3-D-9-glucoside of 55-20a-amino-3 8-hydroxypregnene, and was reported to possess marked hypotensive properties. The presence of steroid alkaloids in the Tabernaemontaneae was hitherto unknown and it was suggested by Raffauf & Flagler (1960) and Bisset (1961) that the plant material was open to further botanical confir- mation. The roots of the conopharyngia species are used in West Africa to treat fever (Kennedy, 1936), including that of malaria (Watt & Breyer-Brandwijk, 1962). The only report on the pharmacology of conopharyngine is that of Zetler (1964), who included it in a study of some of the effects of 23 natural and semi-synthetic alkaloids 174 P. -

The Geography of Welfare in Benin, Burkina Faso, Côte D'ivoire, and Togo
Public Disclosure Authorized Public Disclosure Authorized The Geography of Welfare in Benin, Burkina Faso, Côte d’Ivoire, and Togo Public Disclosure Authorized Nga Thi Viet Nguyen and Felipe F. Dizon Public Disclosure Authorized 00000_CVR_English.indd 1 12/6/17 2:29 PM November 2017 The Geography of Welfare in Benin, Burkina Faso, Côte d’Ivoire, and Togo Nga Thi Viet Nguyen and Felipe F. Dizon 00000_Geography_Welfare-English.indd 1 11/29/17 3:34 PM Photo Credits Cover page (top): © Georges Tadonki Cover page (center): © Curt Carnemark/World Bank Cover page (bottom): © Curt Carnemark/World Bank Page 1: © Adrian Turner/Flickr Page 7: © Arne Hoel/World Bank Page 15: © Adrian Turner/Flickr Page 32: © Dominic Chavez/World Bank Page 48: © Arne Hoel/World Bank Page 56: © Ami Vitale/World Bank 00000_Geography_Welfare-English.indd 2 12/6/17 3:27 PM Acknowledgments This study was prepared by Nga Thi Viet Nguyen The team greatly benefited from the valuable and Felipe F. Dizon. Additional contributions were support and feedback of Félicien Accrombessy, made by Brian Blankespoor, Michael Norton, and Prosper R. Backiny-Yetna, Roy Katayama, Rose Irvin Rojas. Marina Tolchinsky provided valuable Mungai, and Kané Youssouf. The team also thanks research assistance. Administrative support by Erick Herman Abiassi, Kathleen Beegle, Benjamin Siele Shifferaw Ketema is gratefully acknowledged. Billard, Luc Christiaensen, Quy-Toan Do, Kristen Himelein, Johannes Hoogeveen, Aparajita Goyal, Overall guidance for this report was received from Jacques Morisset, Elisée Ouedraogo, and Ashesh Andrew L. Dabalen. Prasann for their discussion and comments. Joanne Gaskell, Ayah Mahgoub, and Aly Sanoh pro- vided detailed and careful peer review comments. -

State of Anarchy Rebellion and Abuses Against Civilians
September 2007 Volume 19, No. 14(A) State of Anarchy Rebellion and Abuses against Civilians Executive Summary.................................................................................................. 1 The APRD Rebellion............................................................................................ 6 The UFDR Rebellion............................................................................................ 6 Abuses by FACA and GP Forces........................................................................... 6 Rebel Abuses....................................................................................................10 The Need for Protection..................................................................................... 12 The Need for Accountability .............................................................................. 12 Glossary.................................................................................................................18 Maps of Central African Republic ...........................................................................20 Recommendations .................................................................................................22 To the Government of the Central African Republic ............................................22 To the APRD, UFDR and other rebel factions.......................................................22 To the Government of Chad...............................................................................22 To the United Nations Security -

Central African Republic: Floods
DREF operation n° MDRCF007 Central African Republic: GLIDE n° FL-2010-000168-CAF Floods 26 August, 2010 The International Federation of Red Cross and Red Crescent (IFRC) Disaster Relief Emergency Fund (DREF) is a source of un-earmarked money created by the Federation in 1985 to ensure that immediate financial support is available for Red Cross and Red Crescent emergency response. The DREF is a vital part of the International Federation’s disaster response system and increases the ability of National Societies to respond to disasters. CHF 145,252 (USD 137,758 or EUR 104,784) has been allocated from the IFRC’s Disaster Relief Emergency Fund (DREF) to support the Central African Red Cross Society (CARCS) in delivering immediate assistance to some 330 displaced families, i.e. 1,650 beneficiaries. Un- earmarked funds to repay DREF are encouraged. Summary: The rainy season that started in July in Central African Republic (CAR) reached its peak on 7 August, 2010 when torrential rains caused serious floods in Bossangoa, a Northern Evaluation of the situation in Bossangoa by CAR Red locality situated 350 km from Bangui, the capital Cross volunteers / Danielle L. Ngaissio, CAR Red Cross city. Other neighbouring localities such as Nanga- Boguila and Kombe, located 187 and 20 km respectively from Bossangoa have also been affected. Damages registered include the destruction of 587 houses (330 completely), the destruction of 1,312 latrines, the contamination of 531 water wells by rain waters and the content of the latrines that have been destroyed. About 2,598 people (505 families) have been affected by the floods. -

Central African Republic: Who Has a Sub-Office/Base Where? (05 May 2014)
Central African Republic: Who has a Sub-Office/Base where? (05 May 2014) LEGEND DRC IRC DRC Sub-office or base location Coopi MSF-E SCI MSF-E SUDAN DRC Solidarités ICRC ICDI United Nations Agency PU-AMI MENTOR CRCA TGH DRC LWF Red Cross and Red Crescent MSF-F MENTOR OCHA IMC Movement ICRC Birao CRCA UNHCR ICRC MSF-E CRCA International Non-Governmental OCHA UNICEF Organization (NGO) Sikikédé UNHCR CHAD WFP ACF IMC UNDSS UNDSS Tiringoulou CRS TGH WFP UNFPA ICRC Coopi MFS-H WHO Ouanda-Djallé MSF-H DRC IMC SFCG SOUTH FCA DRC Ndélé IMC SUDAN IRC Sam-Ouandja War Child MSF-F SOS VdE Ouadda Coopi Coopi CRCA Ngaounday IMC Markounda Kabo ICRC OCHA MSF-F UNHCR Paoua Batangafo Kaga-Bandoro Koui Boguila UNICEF Bocaranga TGH Coopi Mbrès Bria WFP Bouca SCI CRS INVISIBLE FAO Bossangoa MSF-H CHILDREN UNDSS Bozoum COHEB Grimari Bakouma SCI UNFPA Sibut Bambari Bouar SFCG Yaloké Mboki ACTED Bossembélé ICRC MSF-F ACF Obo Cordaid Alindao Zémio CRCA SCI Rafaï MSF-F Bangassou Carnot ACTED Cordaid Bangui* ALIMA ACTED Berbérati Boda Mobaye Coopi CRS Coopi DRC Bimbo EMERGENCY Ouango COHEB Mercy Corps Mercy Corps CRS FCA Mbaïki ACF Cordaid SCI SCI IMC Batalimo CRS Mercy Corps TGH MSF-H Nola COHEB Mercy Corps SFCG MSF-CH IMC SFCG COOPI SCI MSF-B ICRC SCI MSF-H ICRC ICDI CRS SCI CRCA ACF COOPI ICRC UNHCR IMC AHA WFP UNHCR AHA CRF UNDSS MSF-CH OIM UNDSS COHEB OCHA WFP FAO ACTED DEMOCRATIC WHO PU-AMI UNHCR UNDSS WHO CRF MSF-H MSF-B UNFPA REPUBLIC UNICEF UNICEF 50km *More than 50 humanitarian organizations work in the CAR with an office in Bangui. -
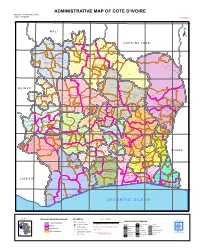
ADMINISTRATIVE MAP of COTE D'ivoire Map Nº: 01-000-June-2005 COTE D'ivoire 2Nd Edition
ADMINISTRATIVE MAP OF COTE D'IVOIRE Map Nº: 01-000-June-2005 COTE D'IVOIRE 2nd Edition 8°0'0"W 7°0'0"W 6°0'0"W 5°0'0"W 4°0'0"W 3°0'0"W 11°0'0"N 11°0'0"N M A L I Papara Débété ! !. Zanasso ! Diamankani ! TENGRELA [! ± San Koronani Kimbirila-Nord ! Toumoukoro Kanakono ! ! ! ! ! !. Ouelli Lomara Ouamélhoro Bolona ! ! Mahandiana-Sokourani Tienko ! ! B U R K I N A F A S O !. Kouban Bougou ! Blésségué ! Sokoro ! Niéllé Tahara Tiogo !. ! ! Katogo Mahalé ! ! ! Solognougo Ouara Diawala Tienny ! Tiorotiérié ! ! !. Kaouara Sananférédougou ! ! Sanhala Sandrégué Nambingué Goulia ! ! ! 10°0'0"N Tindara Minigan !. ! Kaloa !. ! M'Bengué N'dénou !. ! Ouangolodougou 10°0'0"N !. ! Tounvré Baya Fengolo ! ! Poungbé !. Kouto ! Samantiguila Kaniasso Monogo Nakélé ! ! Mamougoula ! !. !. ! Manadoun Kouroumba !.Gbon !.Kasséré Katiali ! ! ! !. Banankoro ! Landiougou Pitiengomon Doropo Dabadougou-Mafélé !. Kolia ! Tougbo Gogo ! Kimbirila Sud Nambonkaha ! ! ! ! Dembasso ! Tiasso DENGUELE REGION ! Samango ! SAVANES REGION ! ! Danoa Ngoloblasso Fononvogo ! Siansoba Taoura ! SODEFEL Varalé ! Nganon ! ! ! Madiani Niofouin Niofouin Gbéléban !. !. Village A Nyamoin !. Dabadougou Sinémentiali ! FERKESSEDOUGOU Téhini ! ! Koni ! Lafokpokaha !. Angai Tiémé ! ! [! Ouango-Fitini ! Lataha !. Village B ! !. Bodonon ! ! Seydougou ODIENNE BOUNDIALI Ponondougou Nangakaha ! ! Sokoro 1 Kokoun [! ! ! M'bengué-Bougou !. ! Séguétiélé ! Nangoukaha Balékaha /" Siempurgo ! ! Village C !. ! ! Koumbala Lingoho ! Bouko Koumbolokoro Nazinékaha Kounzié ! ! KORHOGO Nongotiénékaha Togoniéré ! Sirana -

Central African Rep.: Sub-Prefectures 09 Jun 2015
Central African Rep.: Sub-Prefectures 09 Jun 2015 NIGERIA Maroua SUDAN Birao Birao Abyei REP. OF Garoua CHAD Ouanda-Djallé Ouanda-Djalle Ndélé Ndele Ouadda Ouadda Kabo Bamingui SOUTH Markounda Kabo Ngaounday Bamingui SUDAN Markounda CAMEROON Djakon Mbodo Dompta Batangafo Yalinga Goundjel Ndip Ngaoundaye Boguila Batangafo Belel Yamba Paoua Nangha Kaga-Bandoro Digou Bocaranga Nana-Bakassa Borgop Yarmbang Boguila Mbrès Nyambaka Adamou Djohong Ouro-Adde Koui Nana-Bakassa Kaga-Bandoro Dakere Babongo Ngaoui Koui Mboula Mbarang Fada Djohong Garga Pela Bocaranga MbrÞs Bria Djéma Ngam Bigoro Garga Bria Meiganga Alhamdou Bouca Bakala Ippy Yalinga Simi Libona Ngazi Meidougou Bagodo Bozoum Dekoa Goro Ippy Dir Kounde Gadi Lokoti Bozoum Bouca Gbatoua Gbatoua Bakala Foulbe Dékoa Godole Mala Mbale Bossangoa Djema Bindiba Dang Mbonga Bouar Gado Bossemtélé Rafai Patou Garoua-BoulaiBadzere Baboua Bouar Mborguene Baoro Sibut Grimari Bambari Bakouma Yokosire Baboua Bossemptele Sibut Grimari Betare Mombal Bogangolo Bambari Ndokayo Nandoungue Yaloké Bakouma Oya Zémio Sodenou Zembe Baoro Bogangolo Obo Bambouti Ndanga Abba Yaloke Obo Borongo Bossembele Ndjoukou Bambouti Woumbou Mingala Gandima Garga Abba Bossembélé Djoukou Guiwa Sarali Ouli Tocktoyo Mingala Kouango Alindao Yangamo Carnot Damara Kouango Bangassou Rafa´ Zemio Zémio Samba Kette Gadzi Boali Damara Alindao Roma Carnot Boulembe Mboumama Bedobo Amada-Gaza Gadzi Bangassou Adinkol Boubara Amada-Gaza Boganangone Boali Gambo Mandjou Boganangone Kembe Gbakim Gamboula Zangba Gambo Belebina Bombe Kembé Ouango -

5 Geology and Groundwater 5 Geology and Groundwater
5 GEOLOGY AND GROUNDWATER 5 GEOLOGY AND GROUNDWATER Table of Contents Page CHAPTER 1 PRESENT CONDITIONS OF TOPOGRAPHY, GEOLOGY AND HYDROGEOLOGY.................................................................... 5 – 1 1.1 Topography............................................................................................................... 5 – 1 1.2 Geology.................................................................................................................... 5 – 2 1.3 Hydrogeology and Groundwater.............................................................................. 5 – 4 CHAPTER 2 GROUNDWATER RESOURCES POTENTIAL ............................... 5 – 13 2.1 Mechanism of Recharge and Flow of Groundwater ................................................ 5 – 13 2.2 Method for Potential Estimate of Groundwater ....................................................... 5 – 13 2.3 Groundwater Potential ............................................................................................. 5 – 16 2.4 Consideration to Select Priority Area for Groundwater Development Project ........ 5 – 18 CHAPTER 3 GROUNDWATER BALANCE STUDY .............................................. 5 – 21 3.1 Mathod of Groundwater Balance Analysis .............................................................. 5 – 21 3.2 Actual Groundwater Balance in 1998 ...................................................................... 5 – 23 3.3 Future Groundwater Balance in 2015 ...................................................................... 5 – 24 CHAPTER -

International Activity Report 2017
INTERNATIONAL ACTIVITY REPORT 2017 www.msf.org THE MÉDECINS SANS FRONTIÈRES CHARTER Médecins Sans Frontières is a private international association. The association is made up mainly of doctors and health sector workers, and is also open to all other professions which might help in achieving its aims. All of its members agree to honour the following principles: Médecins Sans Frontières provides assistance to populations in distress, to victims of natural or man-made disasters and to victims of armed conflict. They do so irrespective of race, religion, creed or political convictions. Médecins Sans Frontières observes neutrality and impartiality in the name of universal medical ethics and the right to humanitarian assistance, and claims full and unhindered freedom in the exercise of its functions. Members undertake to respect their professional code of ethics and to maintain complete independence from all political, economic or religious powers. As volunteers, members understand the risks and dangers of the missions they carry out and make no claim for themselves or their assigns for any form of compensation other than that which the association might be able to afford them. The country texts in this report provide descriptive overviews of MSF’s operational activities throughout the world between January and December 2017. Staffing figures represent the total full-time equivalent employees per country across the 12 months, for the purposes of comparisons. Country summaries are representational and, owing to space considerations, may not be comprehensive. For more information on our activities in other languages, please visit one of the websites listed on p. 100. The place names and boundaries used in this report do not reflect any position by MSF on their legal status. -

Phylogeny and Systematics of the Rauvolfioideae
PHYLOGENY AND SYSTEMATICS Andre´ O. Simo˜es,2 Tatyana Livshultz,3 Elena OF THE RAUVOLFIOIDEAE Conti,2 and Mary E. Endress2 (APOCYNACEAE) BASED ON MOLECULAR AND MORPHOLOGICAL EVIDENCE1 ABSTRACT To elucidate deeper relationships within Rauvolfioideae (Apocynaceae), a phylogenetic analysis was conducted using sequences from five DNA regions of the chloroplast genome (matK, rbcL, rpl16 intron, rps16 intron, and 39 trnK intron), as well as morphology. Bayesian and parsimony analyses were performed on sequences from 50 taxa of Rauvolfioideae and 16 taxa from Apocynoideae. Neither subfamily is monophyletic, Rauvolfioideae because it is a grade and Apocynoideae because the subfamilies Periplocoideae, Secamonoideae, and Asclepiadoideae nest within it. In addition, three of the nine currently recognized tribes of Rauvolfioideae (Alstonieae, Melodineae, and Vinceae) are polyphyletic. We discuss morphological characters and identify pervasive homoplasy, particularly among fruit and seed characters previously used to delimit tribes in Rauvolfioideae, as the major source of incongruence between traditional classifications and our phylogenetic results. Based on our phylogeny, simple style-heads, syncarpous ovaries, indehiscent fruits, and winged seeds have evolved in parallel numerous times. A revised classification is offered for the subfamily, its tribes, and inclusive genera. Key words: Apocynaceae, classification, homoplasy, molecular phylogenetics, morphology, Rauvolfioideae, system- atics. During the past decade, phylogenetic studies, (Civeyrel et al., 1998; Civeyrel & Rowe, 2001; Liede especially those employing molecular data, have et al., 2002a, b; Rapini et al., 2003; Meve & Liede, significantly improved our understanding of higher- 2002, 2004; Verhoeven et al., 2003; Liede & Meve, level relationships within Apocynaceae s.l., leading to 2004; Liede-Schumann et al., 2005). the recognition of this family as a strongly supported Despite significant insights gained from studies clade composed of the traditional Apocynaceae s. -

See Full Prospectus
G20 COMPACT WITHAFRICA CÔTE D’IVOIRE Investment Opportunities G20 Compact with Africa 8°W To 7°W 6°W 5°W 4°W 3°W Bamako To MALI Sikasso CÔTE D'IVOIRE COUNTRY CONTEXT Tengrel BURKINA To Bobo Dioulasso FASO To Kampti Minignan Folon CITIES AND TOWNS 10°N é 10°N Bagoué o g DISTRICT CAPITALS a SAVANES B DENGUÉLÉ To REGION CAPITALS Batie Odienné Boundiali Ferkessedougou NATIONAL CAPITAL Korhogo K RIVERS Kabadougou o —growth m Macroeconomic stability B Poro Tchologo Bouna To o o u é MAIN ROADS Beyla To c Bounkani Bole n RAILROADS a 9°N l 9°N averaging 9% over past five years, low B a m DISTRICT BOUNDARIES a d n ZANZAN S a AUTONOMOUS DISTRICT and stable inflation, contained fiscal a B N GUINEA s Hambol s WOROBA BOUNDARIES z a i n Worodougou d M r a Dabakala Bafing a Bere REGION BOUNDARIES r deficit; sustainable debt Touba a o u VALLEE DU BANDAMA é INTERNATIONAL BOUNDARIES Séguéla Mankono Katiola Bondoukou 8°N 8°N Gontougo To To Tanda Wenchi Nzerekore Biankouma Béoumi Bouaké Tonkpi Lac de Gbêke Business friendly investment Mont Nimba Haut-Sassandra Kossou Sakassou M'Bahiakro (1,752 m) Man Vavoua Zuenoula Iffou MONTAGNES To Danane SASSANDRA- Sunyani Guemon Tiebissou Belier Agnibilékrou climate—sustained progress over the MARAHOUE Bocanda LACS Daoukro Bangolo Bouaflé 7°N 7°N Daloa YAMOUSSOUKRO Marahoue last four years as measured by Doing Duekoue o Abengourou b GHANA o YAMOUSSOUKRO Dimbokro L Sinfra Guiglo Bongouanou Indenie- Toulepleu Toumodi N'Zi Djuablin Business, Global Competitiveness, Oumé Cavally Issia Belier To Gôh CÔTE D'IVOIRE Monrovia -

Central African Republic: Population Displacement January 2012
Central African Republic: Population Displacement January 2012 94,386 internally displaced persons (IDPs) in 5,652 the Central African Republic (CAR), where close SUDAN 24,951 65,364 Central to 21,500 were newly displaced in 2012 1,429 African refugees 71,601 returnees from within CAR or Birao neighboring countries 12,820 CHAD 6,880 6,516 Vakaga 19,867 refugees from Sudan and the Democratic Republic of the Congo and 225 11,967 asylum-seekers of varying nationalities reside in Ouanda- the CAR and 152,861 Central African refugees 12,428 Djallé Ndélé are living in neighboring countries 3,827 543 Bamingui- 85,092 Central 7,500 Bangoran African refugees 8,736 1,500 2,525 Kabo 812 Ouadda 5,208 SOUTH SUDAN Markounda Bamingui Haute-Kotto Ngaoundaye 500 3,300 Batangafo Kaga- Haut- Paoua Bandoro Mbomou Nana- Nana-Gribizi Koui Boguila 20 6,736 Bria Bocaranga Ouham Ouham Ouaka 5,517 Djéma 1,033 Central 2,3181,964 5,615 African refugees 3,000 Pendé 3,287 2,074 1,507 128 Bossemtélé Kémo Bambari 1,226 Mbomou 800 Baboua Obo Zémio Ombella M'Poko 1,674 Rafaï Nana-Mambéré 5,564 Bakouma Bambouti CAMEROON 6,978 Basse- Bangassou Kotto Mambéré-Kadéï Bangui Lobaye Returnees Mongoumba Internally displaced persons (IDPs) 1,372 Central Refugees Sangha- African refugees Figures by sub-prefecture Mbaéré Returnee DEMOCRATIC movement REPUBLIC OF THE IDP camp IDP CONGO CONGO Refugee camp Refugee 0 50 100 km Sources: Various sources compiled by OCHA CAR Due to diculty in tracking spontaneous returns, breakdown of refugee returnees and IDP returnees is not available at the sub-prefectural level.