Deoxycytidine-Derivative, in Human Cancer Cells
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Discovery and Development of Seliciclib. How Systems Biology
Journal of Biotechnology 202 (2015) 40–49 Contents lists available at ScienceDirect Journal of Biotechnology j ournal homepage: www.elsevier.com/locate/jbiotec Discovery and development of Seliciclib. How systems biology approaches can lead to better drug performance a b c a a,∗ Hilal S. Khalil , Vanio Mitev , Tatyana Vlaykova , Laura Cavicchi , Nikolai Zhelev a CMCBR, SIMBIOS, School of Science, Engineering and Technology, Abertay University, Dundee DD1 1HG, Scotland, UK b Department of Chemistry and Biochemistry, Medical University of Sofia, 1431 Sofia, Bulgaria c Department of Chemistry and Biochemistry, Medical Faculty, Trakia University, Stara Zagora, Bulgaria a r t i c l e i n f o a b s t r a c t Article history: Seliciclib (R-Roscovitine) was identified as an inhibitor of CDKs and has undergone drug development and Received 10 August 2014 clinical testing as an anticancer agent. In this review, the authors describe the discovery of Seliciclib and Received in revised form 26 February 2015 give a brief summary of the biology of the CDKs Seliciclib inhibits. An overview of the published in vitro Accepted 27 February 2015 and in vivo work supporting the development as an anti-cancer agent, from in vitro experiments to animal Available online 6 March 2015 model studies ending with a summary of the clinical trial results and trials underway is presented. In addition some potential non-oncology applications are explored and the potential mode of action of Keywords: Seliciclib in these areas is described. Finally the authors argue that optimisation of the therapeutic effects Seliciclib of kinase inhibitors such as Seliciclib could be enhanced using a systems biology approach involving Systems biology CDK mathematical modelling of the molecular pathways regulating cell growth and division. -

Targeting Fibrosis in the Duchenne Muscular Dystrophy Mice Model: an Uphill Battle
bioRxiv preprint doi: https://doi.org/10.1101/2021.01.20.427485; this version posted January 21, 2021. The copyright holder for this preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. 1 Title: Targeting fibrosis in the Duchenne Muscular Dystrophy mice model: an uphill battle 2 Marine Theret1#, Marcela Low1#, Lucas Rempel1, Fang Fang Li1, Lin Wei Tung1, Osvaldo 3 Contreras3,4, Chih-Kai Chang1, Andrew Wu1, Hesham Soliman1,2, Fabio M.V. Rossi1 4 1School of Biomedical Engineering and the Biomedical Research Centre, Department of Medical 5 Genetics, 2222 Health Sciences Mall, Vancouver, BC, V6T 1Z3, Canada 6 2Department of Pharmacology and Toxicology, Faculty of Pharmaceutical Sciences, Minia 7 University, Minia, Egypt 8 3Developmental and Stem Cell Biology Division, Victor Chang Cardiac Research Institute, 9 Darlinghurst, NSW, 2010, Australia 10 4Departamento de Biología Celular y Molecular and Center for Aging and Regeneration (CARE- 11 ChileUC), Facultad de Ciencias Biológicas, Pontificia Universidad Católica de Chile, 8331150 12 Santiago, Chile 13 # Denotes Co-first authorship 14 15 Keywords: drug screening, fibro/adipogenic progenitors, fibrosis, repair, skeletal muscle. 16 Correspondence to: 17 Marine Theret 18 School of Biomedical Engineering and the Biomedical Research Centre 19 University of British Columbia 20 2222 Health Sciences Mall, Vancouver, British Columbia 21 Tel: +1(604) 822 0441 fax: +1(604) 822 7815 22 Email: [email protected] 1 bioRxiv preprint doi: https://doi.org/10.1101/2021.01.20.427485; this version posted January 21, 2021. The copyright holder for this preprint (which was not certified by peer review) is the author/funder. -
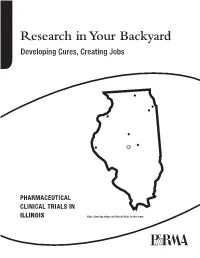
Research in Your Backyard Developing Cures, Creating Jobs
Research in Your Backyard Developing Cures, Creating Jobs PHARMACEUTICAL CLINICAL TRIALS IN ILLINOIS Dots show locations of clinical trials in the state. Executive Summary This report shows that biopharmaceutical research com- Quite often, biopharmaceutical companies hire local panies continue to be vitally important to the economy research institutions to conduct the tests and in Illinois, and patient health in Illinois, despite the recession. they help to bolster local economies in communities all over the state, including Chicago, Decatur, Joliet, Peoria, At a time when the state still faces significant economic Quincy, Rock Island, Rockford and Springfield. challenges, biopharmaceutical research companies are conducting or have conducted more than 4,300 clinical For patients, the trials offer another potential therapeutic trials of new medicines in collaboration with the state’s option. Clinical tests may provide a new avenue of care for clinical research centers, university medical schools and some chronic disease sufferers who are still searching for hospitals. Of the more than 4,300 clinical trials, 2,334 the medicines that are best for them. More than 470 of the target or have targeted the nation’s six most debilitating trials underway in Illinois are still recruiting patients. chronic diseases—asthma, cancer, diabetes, heart dis- ease, mental illnesses and stroke. Participants in clinical trials can: What are Clinical Trials? • Play an active role in their health care. • Gain access to new research treatments before they In the development of new medicines, clinical trials are are widely available. conducted to prove therapeutic safety and effectiveness and compile the evidence needed for the Food and Drug • Obtain expert medical care at leading health care Administration to approve treatments. -

Two Inhibitors of Yeast Plasma Membrane Atpase 1 (Scpma1p): Toward the Development of Novel Antifungal Therapies Sabine Ottilie1†, Gregory M
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by D-Scholarship@Pitt Ottilie et al. J Cheminform (2018) 10:6 https://doi.org/10.1186/s13321-018-0261-3 RESEARCH ARTICLE Open Access Two inhibitors of yeast plasma membrane ATPase 1 (ScPma1p): toward the development of novel antifungal therapies Sabine Ottilie1†, Gregory M. Goldgof1,4†, Andrea L. Cheung1, Jennifer L. Walker2, Edgar Vigil1, Kenneth E. Allen3, Yevgeniya Antonova‑Koch1, Carolyn W. Slayman3^, Yo Suzuki4 and Jacob D. Durrant2* Abstract Given that many antifungal medications are susceptible to evolved resistance, there is a need for novel drugs with unique mechanisms of action. Inhibiting the essential proton pump Pma1p, a P-type ATPase, is a potentially efective therapeutic approach that is orthogonal to existing treatments. We identify NSC11668 and hitachimycin as structur‑ ally distinct antifungals that inhibit yeast ScPma1p. These compounds provide new opportunities for drug discovery aimed at this important target. Keywords: Antifungal, PMA1, P-type ATPase, Computer modeling, Saccharomyces cerevisiae, In vitro evolution, Drug resistance Background sterol-C-24-methyltransferase and the fungal cell mem- Antifungal medications are in high demand, but low brane directly [8]. efcacy, host toxicity, and emerging resistance among Only a few approved antimycotics have mecha- clinical strains [1, 2] complicate their use. Tere is an nisms that are unrelated to ergosterol biosynthesis. urgent need for novel antimycotic therapeutics with For example, the highly efective echinocandins inhibit unique mechanisms of action. Te purpose of the cur- 1,3-β-glucan synthase, hindering production of the criti- rent work is to describe two novel antifungals: 4-N,6- cal cell-wall component β-glucan [9, 10]; and the terato- N-bis(3-chlorophenyl)-1-methylpyrazolo[3,4-d] genic compound fucytosine interferes with eukaryotic pyrimidine-4,6-diamine (NSC11668), and hitachimycin RNA/DNA synthesis [11, 12]. -

Chemotherapeutic Treatment of Acute Myeloid Leukemia
Clinical Trial Outcomes Mahdi, Mahdi & Knapper Chemotherapeutic treatment of acute myeloid leukemia 5 Clinical Trial Outcomes Chemotherapeutic treatment of acute myeloid leukemia: incorporating the results of recent clinical trials Clin. Invest. (Lond.) Better outcomes for patients with acute myeloid leukemia over the last 30 years have Ali J Mahdi*,1, Eamon J been largely achieved by improvements in supportive care measures rather than Mahdi1 & Steven Knapper1 therapeutic advances. The combination of daunorubicin and cytarabine has remained 1Cardiff University School of Medicine, Hematology Department, Heath Park, the standard of care for patients undergoing intensive induction–consolidation Cardiff CF14 4XN, UK treatment. In less fit older patients, low-dose cytarabine is the equivalent, although *Author for correspondence: the hypomethylating agent azacitidine may be challenging current practice. [email protected] Enhanced understanding of disease pathogenesis and therapy resistance has enabled the entry of novel chemotherapeutic and nonchemotherapeutic agents into clinical development with varied levels of activity. This article examines the evidence behind established chemotherapy practices for intensive and nonintensive acute myeloid leukemia treatments with an emphasis on emerging clinical trial data from novel chemotherapeutic and nonchemotherapeutic agents. Keywords: acute myeloid leukemia • aminopeptidase inhibitors • farnesyl transferase inhibitors • FLT3 inhibitors • gemtuzumab ozogamicin • histone deacytylase inhibitors • hypomethylating • Polo-like kinase 1 inhibitors Methodology AML cases [7]. Older patients over the age of 10.4155/CLI.14.112 An electronic database search (EMBASE, 60 years thus represent an important popula- MEDLINE and PubMed) was undertaken tion from both demographic and therapeutic to identify and review clinical studies in perspectives. Older patients frequently have acute myeloid leukemia (AML) undertaken multiple comorbidities, unfavorable cyto- between 1 January 2009 and 1 June 2014. -

Roscovitine-Treated Hela Cells Finalize Autophagy Later Than Apoptosis by Downregulating Bcl-2
1968 MOLECULAR MEDICINE REPORTS 11: 1968-1974, 2015 Roscovitine-treated HeLa cells finalize autophagy later than apoptosis by downregulating Bcl-2 AJDA COKER-GURKAN1, ELIF DAMLA ARISAN1, PINAR OBAKAN1, PELIN OZFILIZ1, BETSI KOSE1, GUVEN BICKICI1,2 and NARCIN PALAVAN-UNSAL1 1Department of Molecular Biology and Genetics, Istanbul Kultur University, Istanbul 34156, Turkey; 2Department of Life and Sport Sciences, School of Science, University of Greenwich, Kent, UK Received December 4, 2013; Accepted May 30, 2014 DOI: 10.3892/mmr.2014.2902 Abstract. The cell cycle is tightly regulated by the family of by roscovitine treatment. The expression levels of different cyclin-dependent kinases (CDKs). CDKs act as regulatory Bcl-2 family members determined whether apoptosis or factors on serine and threonine residues by phosphorylating autophagy were induced following incubation with roscovitine their substrates and cyclins. CDK-targeting drugs have previ- for different time periods. Downregulation of pro-apoptotic ously demonstrated promising effects as cancer therapeutics Bcl-2 family members indicated induction of apoptosis, while both in vitro and in vivo. Roscovitine, a purine-derivative the downregulation of anti-apoptotic Bcl-2 family members and specific CDK inhibitor, has been demonstrated to arrest rapidly induced autophagosome formation in HeLa cells. the cell cycle and induce apoptosis in a number of different cancer cell lines, including HeLa cervical cancer cells. In the Introduction present study, roscovitine was able to decrease both the cell viability and cell survival as well as induce apoptosis in a Cyclin-dependent kinases (CDKs) strictly orchestrate the cell dose-dependent manner in HeLa cells by modulating the mito- cycle machinery through the binding to their specific cyclin chondrial membrane potential. -

Draft COMP Agenda 16-18 January 2018
12 January 2018 EMA/COMP/818236/2017 Inspections, Human Medicines Pharmacovigilance and Committees Committee for Orphan Medicinal Products (COMP) Draft agenda for the meeting on 16-18 January 2018 Chair: Bruno Sepodes – Vice-Chair: Lesley Greene 16 January 2018, 09:00-19:30, room 2F 17 January 2018, 08:30-19:30, room 2F 18 January 2018, 08:30-18:30, room 2F Health and safety information In accordance with the Agency’s health and safety policy, delegates are to be briefed on health, safety and emergency information and procedures prior to the start of the meeting. Disclaimers Some of the information contained in this agenda is considered commercially confidential or sensitive and therefore not disclosed. With regard to intended therapeutic indications or procedure scopes listed against products, it must be noted that these may not reflect the full wording proposed by applicants and may also vary during the course of the review. Additional details on some of these procedures will be published in the COMP meeting reports once the procedures are finalised. Of note, this agenda is a working document primarily designed for COMP members and the work the Committee undertakes. Note on access to documents Some documents mentioned in the agenda cannot be released at present following a request for access to documents within the framework of Regulation (EC) No 1049/2001 as they are subject to on- going procedures for which a final decision has not yet been adopted. They will become public when adopted or considered public according to the principles stated in the Agency policy on access to documents (EMA/127362/2006). -

WO 2010/135468 Al
(12) INTERNATIONAL APPLICATION PUBLISHED UNDER THE PATENT COOPERATION TREATY (PCT) (19) World Intellectual Property Organization International Bureau (10) International Publication Number (43) International Publication Date 25 November 2010 (25.11.2010) WO 2010/135468 Al (51) International Patent Classification: [US/ES]; C/ Rio EsIa 77 PO2 A Villamayor, E-37007 GOlN 33/50 (2006.01) Salamanca (ES). LAGO, Santiago [GB/ES]; C/ Fernando Camino 15 3-E, E-29016 Malaga (ES). MATOSES, (21) International Application Number: Maria [ES/ES]; C/ Los Cristos 1 1 C, E-29008 Malaga PCT/US2010/035474 (ES). SUAREZ, Lilia [ES/ES]; C/ Ancla, 27 Casa 13, (22) International Filing Date: E-29720 CaIa Del Moral (ES). SAPIA, Sandra [IT/ES]; 19 May 2010 (19.05.2010) C/ Zapateros, 2 2 C, E-29005 Malaga (ES). BOSAN- QUET, Andrew [GB/GB]; 4 The Brow, Church Road, English (25) Filing Language: Combe Down, BA2 5JL, Bath (GB). GOR- (26) Publication Language: English ROCHATEGUI, Julian [ES/ES]; C/Puerto de los Leones 5, Majadahonda, E-28220 Madrid (ES). (30) Priority Data: TUDELA, Consuelo [ES/ES]; Av. Juan Carlos I 23, M a 61/179,685 19 May 2009 (19.05.2009) US jadahonda, E-28220 Madrid (ES). HERNANDEZ, Pilar (71) Applicant (for all designated States except US): VIVIA [ES/ES]; C/Uruguay 16, E-37OO3 Salamanca (ES). BIOTECH S.L. [ES/ES]; C/ Menendez Pelayo 12, 3d, CAVEDA, Luis Ignacio [ES/ES]; Av. Valladolid 17, E-47001 Valladolid (ES). E-28008 Madrid (ES). (72) Inventors; and (74) Agent: MALLON, Joseph, J.; Knobbe Martens Olson & (75) Inventors/Applicants (for US only): BALLESTEROS, Bear LLP, 2040 Main Street, 14th Floor, Irvine, CA Juan [ES/ES]; C/ Espalter 8 - 7 B, E-28014 Madrid (ES). -

Peptide-Functionalized Nanoparticles-Encapsulated Cyclin-Dependent Kinases Inhibitor Seliciclib in Transferrin Receptor Overexpressed Cancer Cells
nanomaterials Article Peptide-Functionalized Nanoparticles-Encapsulated Cyclin-Dependent Kinases Inhibitor Seliciclib in Transferrin Receptor Overexpressed Cancer Cells Guan Zhen He 1 and Wen Jen Lin 1,2,* 1 School of Pharmacy, College of Medicine, National Taiwan University, Taipei 10050, Taiwan; [email protected] 2 Drug Research Center, College of Medicine, National Taiwan University, Taipei 10050, Taiwan * Correspondence: [email protected]; Tel.: +886-2-33668765; Fax: +886-2-23919098 Abstract: Seliciclib, a broad cyclin-dependent kinases (CDKs) inhibitor, exerts its potential role in cancer therapy. For taking advantage of overexpressive transferrin receptor (TfR) on most cancer cells, T7 peptide, a TfR targeting ligand, was selected as a targeting ligand to facilitate nanoparticles (NPs) internalization in cancer cells. In this study, poly(D,L-lactide-co-glycolide) (PLGA) was conju- gated with maleimide poly(ethylene glycol) amine (Mal-PEG-NH2) to form PLGA-PEG-maleimide copolymer. The synthesized copolymer was used to prepare NPs for encapsulation of seliciclib which was further decorated by T7 peptide. The result shows that the better cellular uptake was achieved by T7 peptide-modified NPs particularly in TfR-high expressed cancer cells in order of MDA-MB-231 breast cancer cells > SKOV-3 ovarian cancer cells > U87-MG glioma cells. Both SKOV-3 and U87-MG cells are more sensitive to encapsulated seliciclib in T7-decorated NPs than to free seliciclib, and that Citation: He, G.Z.; Lin, W.J. IC50 values were lowered for encapsulated seliciclib. Peptide-Functionalized Nanoparticles-Encapsulated Keywords: seliciclib; T7 peptide; nanoparticles; TfR-overexpressed cancer cells Cyclin-Dependent Kinases Inhibitor Seliciclib in Transferrin Receptor Overexpressed Cancer Cells. -

Roscovitine in Cancer and Other Diseases
Review Article Page 1 of 12 Roscovitine in cancer and other diseases Jonas Cicenas1,2,3, Karthik Kalyan2,4, Aleksandras Sorokinas2, Edvinas Stankunas2,5, Josh Levy2,6, Ingrida Meskinyte7, Vaidotas Stankevicius2,8,9, Algirdas Kaupinis3, Mindaugas Valius3 1CALIPHO Group, Swiss Institute of Bioinformatics, Geneva, Switzerland; 2MAP Kinase Resource, Bern, Switzerland; 3Proteomics Centre, Vilnius University Institute of Biochemistry, Vilnius, Lithuania; 4Systems Biomedicine Division and Department of Virology and Immunology, Haffkine Institute for Training Research and Testing, Mumbai, India; 5Department of Biochemistry, Vilnius University, Vilnius, Lithuania; 6RTI International, Research Triangle Park, NC, USA; 7Lithuanian Centre of Non-Formal Youth Education Vilnius, Lithuania; 8National Cancer Institute, Vilnius, Lithuania; 9Vilnius University, Vilnius, Lithuania Correspondence to: Jonas Cicenas. Swiss Institute of Bioinformatics, CALIPHO Group, CMU-1, rue Michel Servet’ CH-1211, Geneva 4, Switzerland. Email: [email protected]. Abstract: Roscovitine [CY-202, (R)-Roscovitine, Seliciclib] is a small molecule that inhibits cyclin-dependent kinases (CDKs) through direct competition at the ATP-binding site. It is a broad-range purine inhibitor, which inhibits CDK1, CDK2, CDK5 and CDK7, but is a poor inhibitor for CDK4 and CDK6. Roscovitine is widely used as a biological tool in cell cycle, cancer, apoptosis and neurobiology studies. Moreover, it is currently evaluated as a potential drug to treat cancers, neurodegenerative diseases, inflammation, viral infections, polycystic kidney disease and glomerulonephritis. This review focuses on the use of roscovitine in the disease model as well as clinical model research. Keywords: Cyclin-dependent kinases (CDK); small molecule inhibitor; roscovitine; cancer; neurodegeneration; kidney diseases Submitted Dec 16, 2014. Accepted for publication Mar 16, 2015. -

20 13 Rep Or T
2013 REPORT More Than 240 Medicines in Development for Leukemia, Lymphoma and Other Blood Cancers Every 4 minutes a person is diagnosed with leukemia, Medicines in Development lymphoma or For Leading Blood Cancers myeloma; Accounting for Application Submitted 9% of all cancers Phase III diagnosed each year Phase II Biopharmaceutical research companies • 15 each for myeloproliferative neo- Phase I are developing 241 medicines for blood plasms, such as myelofibrosis, poly- cancers—leukemia, lymphoma and cythemia vera and essential throm- 97 98 myeloma. This report lists medicines in bocythemia; and for myelodysplastic human clinical trials or under review by syndromes, which are diseases affect- the U.S. Food and Drug Administration ing the blood and bone marrow. (FDA). These medicines in development offer The medicines in development include: hope for greater survival for the thou- sands of Americans who are affected by • 98 for lymphoma, including Hodgkin these cancers of the blood. and non-Hodgkin lymphoma, which 52 affect nearly 80,000 Americans each Definitions for the cancers listed in this year. report and other terms can be found on page 27. Links to sponsor company web • 97 for leukemia, including the four sites provide more information on the major types, which affect nearly potential products. 50,000 people in the United States 24 each year. For information on the value of medi- cines, an in-depth look at current in- • 52 for myeloma, a cancer of the novation and key medical breakthroughs plasma cells, which impacts more benefiting blood cancer patients, please than 22,000 people each year in the see Medicines in Development for Leu- United States. -

In Combination with Cytotoxic Agents in Human Uterine Sarcoma Cell Lines
ANTICANCER RESEARCH 27: 273-278 (2007) Seliciclib (CYC202; r-Roscovitine) in Combination with Cytotoxic Agents in Human Uterine Sarcoma Cell Lines HELEN M. COLEY, CHRISTINE F. SHOTTON and HILARY THOMAS Postgraduate Medical School, University of Surrey, Guildford, Surrey GU2 7WG, U.K. Abstract. Background: Inhibition of cyclin-dependent kinases the cell cycle as an approach to treat cancer. Seliciclib (CDKs) has recently emerged as an interesting approach to treat (CYC202), the r-enantiomer of the cell cycle inhibitory human malignancies. This was explored in human leiomyo- agent roscovitine has been developed as a potent CDK2 sarcoma (LMS) lines, which represent a tumour associated with inhibitor and is currently in phase II clinical trials. poor survival, chemo-unresponsiveness and deregulation of cell Preclinical studies involving other CDK inhibitors, such as cycle components. Materials and Methods: Using isobologram flavopiridol have demonstrated their interaction with a analysis with MTT chemosensitivity testing, the effects of the CDK number of different cytotoxic agents in a synergistic manner inhibitor seliciclib (CYC202, R-roscovitine) when used alone or (5). We have explored this approach by examining the in combination with paclitaxel was studied in uterine cancer cell effects of seliciclib combined with paclitaxel in three human lines. Apoptotic endpoints were also examined via Annexin V uterine sarcoma cell line models in terms of any synergy and assay using flow cytometry and Western blotting. Results: Overall effects on apoptosis. seliciclib combined with paclitaxel proved synergistic for all cell lines. This was concomitant with an enhanced apoptotic effect Materials and Methods and downregulation of the IAP survivin. Conclusion: Our data support the use of seliciclib as part of combination therapy for Chemicals and reagents.