Fossils from the Southern Hemisphere
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Outdoor Recreation Strategy 1 2012 - 2022 Central Otago Outdoor Recreation Sstrategytrategy 2012012222 --- 2022022222
= `Éåíê~ä=lí~Öç= =lìíÇççê=oÉÅêÉ~íáçå= píê~íÉÖó= = OMNO=J=OMOO= February 2012 This is a community owned strategy developed by the Outdoor Recreation Working Party in consultation with the Central Otago Community Central Otago Outdoor Recreation Strategy 1 2012 - 2022 Central Otago Outdoor Recreation SStrategytrategy 2012012222 --- 2022022222 PAGE EXECUTIVE SUMMARY 4 IMPLEMENTATION 8 INTRODUCTION 15 Goals 15 Why have an Outdoor Recreation Strategy? 15 What Comprises Recreation? 16 What Makes a Good Experience 16 Purpose 16 Management Approaches 16 Planning 17 Importance of Outdoor Recreation 17 Central Otago – Geographically Defined 17 Barriers to Participation in Outdoor Recreation 18 Changing Perceptions of Outdoor Recreation 19 Fragmentation of Leisure Time 19 Conflict of Use 19 Changing Perceptions of Risk 19 Developing Outdoor Skills 20 Outdoor Recreation, Individuals and Communities 20 Environmental Considerations 21 Economic Considerations 21 Key Characteristics of Central Otago 21 Other Strategies 21 Regional Identity (A World of Difference) 22 Other Agencies and Groups Involved 22 Assumptions and Uncertainties 22 OVERARCHING ISSUES Human Waste Disposal 23 Rubbish 23 Dogs 23 Signs, Route Guides and Waymarking (Geographic Information) 24 Access 24 Research 25 Landowners 25 Competing Use 26 Communications 27 SPECIFIC RECREATION ACTIVITIES Notes on Tracks, Trails and Recreational Areas 28 Air Activities 29 Mountain Biking 31 Road Cycling 38 Climbing 40 Four Wheel Driving 43 Gold Panning 47 Hunting – Small Game and Big Game 49 Central -

§4-71-6.5 LIST of CONDITIONALLY APPROVED ANIMALS November
§4-71-6.5 LIST OF CONDITIONALLY APPROVED ANIMALS November 28, 2006 SCIENTIFIC NAME COMMON NAME INVERTEBRATES PHYLUM Annelida CLASS Oligochaeta ORDER Plesiopora FAMILY Tubificidae Tubifex (all species in genus) worm, tubifex PHYLUM Arthropoda CLASS Crustacea ORDER Anostraca FAMILY Artemiidae Artemia (all species in genus) shrimp, brine ORDER Cladocera FAMILY Daphnidae Daphnia (all species in genus) flea, water ORDER Decapoda FAMILY Atelecyclidae Erimacrus isenbeckii crab, horsehair FAMILY Cancridae Cancer antennarius crab, California rock Cancer anthonyi crab, yellowstone Cancer borealis crab, Jonah Cancer magister crab, dungeness Cancer productus crab, rock (red) FAMILY Geryonidae Geryon affinis crab, golden FAMILY Lithodidae Paralithodes camtschatica crab, Alaskan king FAMILY Majidae Chionocetes bairdi crab, snow Chionocetes opilio crab, snow 1 CONDITIONAL ANIMAL LIST §4-71-6.5 SCIENTIFIC NAME COMMON NAME Chionocetes tanneri crab, snow FAMILY Nephropidae Homarus (all species in genus) lobster, true FAMILY Palaemonidae Macrobrachium lar shrimp, freshwater Macrobrachium rosenbergi prawn, giant long-legged FAMILY Palinuridae Jasus (all species in genus) crayfish, saltwater; lobster Panulirus argus lobster, Atlantic spiny Panulirus longipes femoristriga crayfish, saltwater Panulirus pencillatus lobster, spiny FAMILY Portunidae Callinectes sapidus crab, blue Scylla serrata crab, Samoan; serrate, swimming FAMILY Raninidae Ranina ranina crab, spanner; red frog, Hawaiian CLASS Insecta ORDER Coleoptera FAMILY Tenebrionidae Tenebrio molitor mealworm, -

Natural Character, Riverscape & Visual Amenity Assessments
Natural Character, Riverscape & Visual Amenity Assessments Clutha/Mata-Au Water Quantity Plan Change – Stage 1 Prepared for Otago Regional Council 15 October 2018 Document Quality Assurance Bibliographic reference for citation: Boffa Miskell Limited 2018. Natural Character, Riverscape & Visual Amenity Assessments: Clutha/Mata-Au Water Quantity Plan Change- Stage 1. Report prepared by Boffa Miskell Limited for Otago Regional Council. Prepared by: Bron Faulkner Senior Principal/ Landscape Architect Boffa Miskell Limited Sue McManaway Landscape Architect Landwriters Reviewed by: Yvonne Pfluger Senior Principal / Landscape Planner Boffa Miskell Limited Status: Final Revision / version: B Issue date: 15 October 2018 Use and Reliance This report has been prepared by Boffa Miskell Limited on the specific instructions of our Client. It is solely for our Client’s use for the purpose for which it is intended in accordance with the agreed scope of work. Boffa Miskell does not accept any liability or responsibility in relation to the use of this report contrary to the above, or to any person other than the Client. Any use or reliance by a third party is at that party's own risk. Where information has been supplied by the Client or obtained from other external sources, it has been assumed that it is accurate, without independent verification, unless otherwise indicated. No liability or responsibility is accepted by Boffa Miskell Limited for any errors or omissions to the extent that they arise from inaccurate information provided by the Client or -

Miocene Mammal Reveals a Mesozoic Ghost Lineage on Insular New Zealand, Southwest Pacific
Miocene mammal reveals a Mesozoic ghost lineage on insular New Zealand, southwest Pacific Trevor H. Worthy*†, Alan J. D. Tennyson‡, Michael Archer§, Anne M. Musser¶, Suzanne J. Hand§, Craig Jonesʈ, Barry J. Douglas**, James A. McNamara††, and Robin M. D. Beck§ *School of Earth and Environmental Sciences, Darling Building DP 418, Adelaide University, North Terrace, Adelaide 5005, South Australia, Australia; ‡Museum of New Zealand Te Papa Tongarewa, P.O. Box 467, Wellington 6015, New Zealand; §School of Biological, Earth and Environmental Sciences, University of New South Wales, New South Wales 2052, Australia; ¶Australian Museum, 6-8 College Street, Sydney, New South Wales 2010, Australia; ʈInstitute of Geological and Nuclear Sciences, P.O. Box 30368, Lower Hutt 5040, New Zealand; **Douglas Geological Consultants, 14 Jubilee Street, Dunedin 9011, New Zealand; and ††South Australian Museum, Adelaide, South Australia 5000, Australia Edited by James P. Kennett, University of California, Santa Barbara, CA, and approved October 11, 2006 (sent for review July 8, 2006) New Zealand (NZ) has long been upheld as the archetypical Ma) dinosaur material (13) and isolated moa bones from marine example of a land where the biota evolved without nonvolant sediments up to 2.5 Ma (1, 14), the terrestrial record older than terrestrial mammals. Their absence before human arrival is mys- 1 Ma is extremely limited. Until now, there has been no direct terious, because NZ was still attached to East Antarctica in the Early evidence for the pre-Pleistocene presence in NZ of any of its Cretaceous when a variety of terrestrial mammals occupied the endemic vertebrate lineages, particularly any group of terrestrial adjacent Australian portion of Gondwana. -
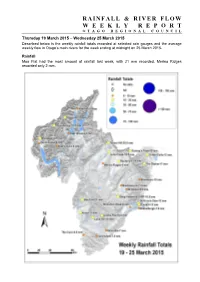
Rainfall & River Flow Weeklyreport
RAINFALL & RIVER FLOW WEEKLY REPORT OTAGO REGIONAL COUNCIL Thursday 19 March 2015 – Wednesday 25 March 2015 Described below is the weekly rainfall totals recorded at selected rain gauges and the average weekly flow in Otago’s main rivers for the week ending at midnight on 25 March 2015. Rainfall Moa Flat had the most amount of rainfall last week, with 21 mm recorded. Merino Ridges recorded only 2 mm. River Flows Flows in the Manuherikia River, Shotover River, Kawarau River, Waipahi River, and the Clutha River at Balclutha were below normal. The Kakanui River at Clifton Falls was the only flow recorder having above normal flows. Table 1. River flow information for Otago’s main rivers (all flows in cumecs, m3/s) Weekly River and Site Name Minimum Maximum State Average Kakanui River at Clifton Falls 1.391 0.919 2.680 above normal Shag River at The Grange 0.219 0.164 0.332 normal Taieri River at Canadian Flat 2.126 1.290 4.409 normal Taieri River at Tiroiti 2.728 2.084 5.377 normal Taieri River at Sutton 3.335 2.469 5.636 normal Taieri River at Outram 5.884 4.823 8.000 normal Clutha River at Balclutha 376.966 294.501 505.056 below normal Waipahi River at Waipahi 0.672 0.542 0.905 below normal Pomahaka River at Burkes Ford 9.157 7.009 12.915 normal Manuherikia River at Ophir 1.869 1.418 2.270 below normal Clutha R. at Cardrona Confluence 229.972 145.042 288.669 normal Kawarau River at Chards Rd 126.705 119.148 135.372 below normal Shotover River at Peat's Hut 10.783 10.122 13.659 below normal Lake Levels Water levels in Lake Hawea and Lake Wakatipu were both well below normal. -

Hydrological Impacts of Irrigated Agriculture in the Manuherikia Catchment, Otago, New Zealand
Journal of Hydrology (NZ) 47 (2): 67-84 2008 © New Zealand Hydrological Society (2008) Hydrological impacts of irrigated agriculture in the Manuherikia catchment, Otago, New Zealand Stefan W. Kienzle1 and Jochen Schmidt2 1 Department of Geography, University of Lethbridge, Alberta, Canada 2 National Institute of Water and Atmospheric Research, P.O. Box 8602, Christchurch, New Zealand. Corresponding author: [email protected] Abstract to losses in the conveyance system, on Water for irrigation is becoming an in farm application losses, increased actual creasingly critical component of New evapotranspiration, and the development of Zealand’s rural economy. Projections of a deeper rooting system compared to natural expanding agricultural sectors indicate vegetation. increased demands for water and a reliable water supply. Hence, it is pertinent to assess Introduction the impacts of water demand for current Water for irrigation is becoming an and future irrigated agriculture on catch increasingly critical component of New ment hydrology. This case study focuses Zealand’s rural economy. More intensive on estimating the hydrological impacts of farming systems are usually accompanied by irrigated agriculture in the Manuherikia a demand for increased water quantity and catchment upstream of Alexandra, Central a reliable water supply. Projections indicate Otago, New Zealand. In order to assess that New Zealand’s dairy, horticulture the impacts of irrigation on streamflow, and viticulture sectors will all expand in five landuse scenarios, including three the future, and it follows that there will be irrigation scenarios with varying efficiencies, growing demands for water for irrigated are investigated with the ACRU agro agriculture (Ministry of Agriculture and hydrological modeling system (Agricultural Forestry, 2004; Parliamentary Commissioner Catchments Research Unit (ACRU), for the Environment, 2004; Doak, 2005). -

Pioneer Park Is
Bantry Street - e c n Pioneer a Wood r t Chopping n ALEXANDRA E Trade Displays n Park o si ces - Pro F Licensed Area o - o d F l & o B a e t v s e ra D Art & Craft ge Entrance i s p Markets l Gates a y Carnival - Area Tarbert Street F o Face Painting o d & B e ve ra ge Main Stage Shannon Street Shannon St John Lost KidsKK Kids Stage BNZ ATM B DESIGN BNZ ATM Toilets Site Playground Tennis Courts Museum Centennial Avenue PRINCIPAL PARTNER: SPONSORS: GRANTS Pioneer Park is DISTRICT COUNCIL ALEXANDRA ROUND THE CLOCK RACE Round the Clock Race teams and individuals traverse the Manuherikia River & scramble up the hillside to the Alexandra Clock Race briefing at 5.15pm, Linger & Die, Rivers St Alexandra. Racing starts 5.45pm. Entry forms www.blossom.co.nz Saturday 26th September For additional info Trevor Breen 021 340 713. 11.45am: ATL Trucks Parade of Trucks Procession Order • CO Vintage Car Club with guests from Living Options Monday 21 September • Dunstan Vintage Machinery Club. Vintage Tractors All Day: Blossom Drive – Follow the Blossom Arrows for the • Central Motor Speedway most scenic Blossom Drive in NZ. Begin at the Alexandra Bridge and head towards Earnscleugh. A round trip of 30km. ARDI RAS • Southland Zephyr and Zodiac Club 6pmM – 8.30 pm: Rivers GStreet, Alexandra Main land FMX • Highlands Motorsport Park 10am – 4pm: Denis Kent Retrospective. Central Stories Display, Fireworks Display, special guest Craig Smith the Museum and Art Gallery. Admission Free. Wonkey Donkey Man, Registration • Anti P Ministry Internationally acclaimed local artist Denis Kent celebrates P: Martin 021 879 849, [email protected] • Alexandra Pipe Band his 80th birthday this year. -

Alexandra | Cromwell Tracks Brochure
OTAGO Welcome to Central Otago Nau mai, haere mai Alexandra and Cromwell townships are good bases from which to Alexandra explore Central Otago, a popular outdoor destination for mountain Further information biking, walking, four-wheel driving, fishing and sharing picnics. Cromwell tracks The vast ‘big sky’ landscape offers a variety of adventures and places Tititea/Mt Aspiring National Park Visitor Centre to explore. 1 Ballantyne Road Central Otago Wanaka 9305 Key PHONE: (03) 443 7660 Mountain bike tracks Walking tracks EMAIL: [email protected] Grade 1: Easiest Walking track www.doc.govt.nz EASIEST Grade 2: Easy Short walk Grade 3: Intermediate Tramping track Grade 4: Advanced Route ADVANCED No dogs No horses 4WD Ski touring Historic site Picnic Horse riding Fishing Swimming Dog walking Hunting Lookout Motorcycling Mountain biking Published by: R174401 Tititea/Mount Aspiring National Park Visitor Centre New Zealand Cycle Trail Ardmore Street, Wanaka PO Box 93, Wanaka 9343 Managed by Department of Conservation Phone: 03 443 7660 Email: [email protected] Managed by Central Otago District Council September 2020 Editing and design: Managed by Cromwell & Districts Te Rōpū Ratonga Auaha, Te Papa Atawhai Promotions Group Creative Services, Department of Conservation This publication is produced using paper sourced from Landmarks well-managed, renewable and legally logged forests. Toyota Kiwi Guardians Front page image photo credit: Bannockburn Sluicings. Photo: C. Babirat Mountain Bikers of Alexandra (MOA) Some quick recreation ideas History Choosing a picnic spot Māori Great picnic spots can be found at Lanes Dam, Alexandra (Aronui Although there were never large numbers of Māori living in this area, Dam), Mitchells Cottage and Bendigo/Logantown. -

Clutha River Catchment Water Quality and Ecosystem Health July 2011 to June 2012
Clutha River catchment Water quality and ecosystem health July 2011 to June 2012 Water quality Summary To assess the current state of water quality, the Otago Regional Council (ORC) All of the sites in the upper monitored 37 river and stream sites in the Clutha catchment between July 2011 and Clutha, had either excellent or June 2012. Most sites were monitored every two months, but five further sites good water quality. (Clutha River/Mata-Au at Luggate, Millers Flat, Balclutha, the Kawarau River, and Water quality in south-west the Shotover River) were monitored monthly by NIWA as part of the National River Water Quality Network (NRWQN). Sites were classified using a water quality index. Otago was poorer, particularly in the Pomahaka River tributaries. The Manuherikia River and the Heriot Burn supported the healthiest macroinvertebrate communities. The Lindis River supported growths of both Didymosphenia geminata and Phormidium. Brown trout were common at all monitoring sites. However eels were absent at sites above the Clutha dams. Water quality index ORC uses a water quality index (WQI) derived from median values of six indicator variables, turbidity, dissolved oxygen (percent saturation), ammonical nitrogen (NH4), nitrite-nitrate nitrogen (NNN), dissolved reactive phosphorus (DRP) and Escherichia coli (E. coli). Median values of the six values are compared with ANZECC (2000) and MfE/MoH (2003) guidelines, enabling classification of water quality into one of the four group Excellent All six values comply with guideline values Good Five median values comply Fair Three or four median values comply Poor Two or fewer median values comply with guideline values Guidelines for nutrients Nutrients Nitrite-nitrate nitrogen (NNN) is a form of nitrogen primarily derived from land drainage. -

Supplementary Information
Supplementary Information Substitution Rate Variation in a Robust Procellariiform Seabird Phylogeny is not Solely Explained by Body Mass, Flight Efficiency, Population Size or Life History Traits Andrea Estandía, R. Terry Chesser, Helen F. James, Max A. Levy, Joan Ferrer Obiol, Vincent Bretagnolle, Jacob González-Solís, Andreanna J. Welch This pdf file includes: Supplementary Information Text Figures S1-S7 SUPPLEMENTARY INFORMATION TEXT Fossil calibrations The fossil record of Procellariiformes is sparse when compared with other bird orders, especially its sister order Sphenisciformes (Ksepka & Clarke 2010, Olson 1985c). There are, however, some fossil Procellariiformes that are both robustly dated and identified and therefore suitable for fossil calibrations. Our justification of these fossils, below, follows best practices described by Parham et al. (2012) where possible. For all calibration points only a minimum age was set with no upper constraint specified, except for the root of the tree. 1. Node between Sphenisciformes/Procellariiformes Minimum age: 60.5 Ma Maximum age: 61.5 Ma Taxon and specimen: Waimanu manneringi (Slack et al. 2006); CM zfa35 (Canterbury Museum, Christchurch, New Zealand), holotype comprising thoracic vertebrae, caudal vertebrae, pelvis, femur, tibiotarsus, and tarsometatarsus. Locality: Basal Waipara Greensand, Waipara River, New Zealand. Phylogenetic justification: Waimanu has been resolved as the basal penguin taxon using morphological data (Slack et al. 2006), as well as combined morphological and molecular datasets (Ksepka et al. 2006, Clarke et al. 2007). Morphological and molecular phylogenies agree on the monophyly of Sphenisciformes and Procellariiformes (Livezey & Zusi 2007, Prum et al. 2015). Waimanu manneringi was previously used by Prum et al. (2015) to calibrate Sphenisiciformes, and see Ksepka & Clarke (2015) for a review of the utility of this fossil as a robust calibration point. -

By Rijksmuseum Van Natuurlijke Historie, Leiden in Preparing The
RESULTS OF A REEXAMINATION OF TYPES AND SPECIMENS OF GOBIOID FISHES, WITH NOTES ON THE FISHFAUNA OF THE SURROUNDINGS OF BATAVIA by Dr. F. P. KOUMANS Rijksmuseum van Natuurlijke Historie, Leiden In preparing the volume of the Gobioidea in M. Weber and L. F. de Beaufort: The Fishes of the Indo-Australian Archipelago, several de- scribed species, collected in the Indo-Australian Archipelago or its surroundings, were not clear to me. Of a number of these the description was distinct enough to see what was meant with such a new species, but there were several species which I could not recognize from their description. Bleeker described a large number of new species, but, unfortunately, several of his descriptions are too vague to recognize the species. So many authors had described several species which proved, after comparison with Bleeker's type specimens or descriptions made after his types, to be either closely allied, or identical with species already described by Bleeker. In order to see whether the described species of authors were synonyms of already described species, or to reexamine the types in order to enlarge the descriptions, I visited several Museums and other Institutions in the United States of N. America, Honolulu, Australia, Philippines, Singapore and British India. During a stay in Batavia, I had the opportunity to make colour sketches of freshly-caught specimens and to go out and collect specimens myself. My visit to the different countries mentioned was made possible by a grant of the "Pieter Langerhuizen Lambertuszoon fonds", endowed by the "Hollandsche Maatschappij der Wetenschappen". During these visits I received great help and friendship of the staff of the Museums and Institutions, for which I am very thankful. -

The Parasite Release Hypothesis and the Success of Invasive Fish in New Zealand
http://researchcommons.waikato.ac.nz/ Research Commons at the University of Waikato Copyright Statement: The digital copy of this thesis is protected by the Copyright Act 1994 (New Zealand). The thesis may be consulted by you, provided you comply with the provisions of the Act and the following conditions of use: Any use you make of these documents or images must be for research or private study purposes only, and you may not make them available to any other person. Authors control the copyright of their thesis. You will recognise the author’s right to be identified as the author of the thesis, and due acknowledgement will be made to the author where appropriate. You will obtain the author’s permission before publishing any material from the thesis. The parasite release hypothesis and the success of invasive fish in New Zealand A thesis submitted in partial fulfilment of the requirements for the degree of Master of Science in Biological Science at The University of Waikato by Keshi Zhang The University of Waikato 2012 Abstract Non-indigenous species are commonly released from their native enemies, including parasites, when they are introduced into new geographical areas. This has been referred to as the enemy release hypothesis and more strictly as the parasite release hypothesis. The loss of parasites is commonly inferred to explain the invasiveness of non-indigenous species. I examined parasite release in New Zealand non-indigenous freshwater fishes. A literature review was undertaken in order to collate lists of the known parasite fauna of 20 New Zealand non-indigenous freshwater fish species.