The Status of Caspian Tern Sterna Caspia
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Beaufort Seas West To
71° 162° 160° 158° 72° U LEGEND 12N 42W Ch u $ North Slope Planning Area ckchi Sea Conservation System Unit (Offset for display) Pingasagruk (abandoned) WAINWRIGHT Atanik (Abandoned) Naval Arctic Research Laboratory USGS 250k Quad Boundaries U Point Barrow I c U Point Belcher 24N Township Boundaries y 72° Akeonik (Ruins) Icy Cape U 17W C 12N Browerville a Trans-Alaska Pipeline p 39W 22N Solivik Island e Akvat !. P Ikpilgok 20W Barrow Secondary Roads (unpaved) a Asiniak!. Point s MEADE RIVER s !. !. Plover Point !. Wainwright Point Franklin !. Brant Point !. Will Rogers and Wiley Post Memorial Whales1 U Point Collie Tolageak (Abandoned) 9N Point Marsh Emaiksoun Lake Kilmantavi (Abandoned) !. Kugrua BayEluksingiak Point Seahorse Islands Bowhead Whale, Major Adult Area (June-September) 42W Kasegaluk Lagoon West Twin TekegakrokLake Point ak Pass Sigeakruk Point uitk A Mitliktavik (Abandoned) Peard Bay l U Ikroavik Lake E Tapkaluk Islands k Wainwright Inlet o P U Bowhead Whale, Major Adult Area (May) l 12N k U i i e a n U re Avak Inlet Avak Point k 36W g C 16N 22N a o Karmuk Point Tutolivik n U Elson Lagoon t r !. u a White (Beluga) Whale, Major Adult Area (September) !. a !. 14N m 29W 17W t r 17N u W Nivat Point o g P 32W Av k a a Nokotlek Point !. 26W Nulavik l A s P v a a s Nalimiut Point k a k White (Beluga) Whale, Major Adult Area (May-September) MEADE RIVER p R s Pingorarok Hill BARROW U a Scott Point i s r Akunik Pass Kugachiak Creek v ve e i !. -

Non-Living (Abiotic) Elements Shape Habitat
Forest Birds Fast Facts LIGHT: Dominated by tall trees, layered canopy = very shady, openings made from a fallen tree provide sunny areas AIR: Trees slow wind, except along forest edges and openings. Shade = cool temps WATER: Fog and rain collect in tree branches and drip to ground. SOIL: Lots of organics (needles, decaying leaves, tree trunks) makes lots of space for air and water. Soil absorbs and holds water like a sponge. Varied Thrush • Slender bill good at gleaning soft foods like insects, pillbugs, snails, worms, fruits and some seeds from ground. • Hops and pauses to look for food. Flips leaves and debris with bill. • Perching feet – three toes forward, hind toe back. • Can sing 2 separate notes at same time and breathe while singing. Common Raven Varied Thrush • Versatile beak. Eats everything from carrion and garbage, to eggs, nestlings, insects, seeds, rodents and fruit. • Strong, sturdy feet and grasping toes to manipulate food and perch. • Acrobatic flight, hops on ground. Uses sight to find food. Northern Flicker • The toes are placed two forward, two back to grip firmly, while the tail feathers are stiff and pointed to help brace while pounding. • Bill is shaped like a chisel. Flickers don’t excavate as much as other types of woodpeckers so have a slightly curved bill and less sharp. Common Raven • Flickers eat insects and are especially fond of ants (flickers will forage on the ground as well as on trees). Probe and explore crevices. • Distinctive flight – flap, dip, flap, dip • Woodpeckers have long, sticky and barbed tongues to extract bugs. -
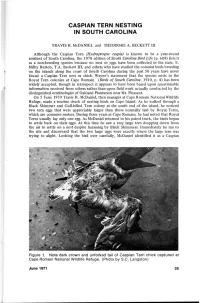
Caspian Tern Nesting in South Carolina
CASPIAN TERN NESTING IN SOUTH CAROLINA TRAVIS H. McDANIEL and THEODORE A. BECKETT III Although the Caspian Tern (Hydroprogne caspia) is known to be a year-round resident of South Carolina, the 1970 edition of South Carolina Bird Life (p. 608) lists it as a non-breeding species because no nest or eggs have been collected in the state. E. Milby Burton, T.A. Beckett III, and others who have studied the colonial birds breeding on the islands along the coast of South Carolina during the past 50 years have never found a Caspian Tern nest or chick. Wayne's statement that the species nests in the Royal Tern colonies at Cape Romain (Birds of South Carolina, 1910, p. 4) has been widely accepted, though in retrospect it appears to have been based upon questionable information received from others rather than upon field work actually conducted by the distinguished ornithologist of Oakland Plantation near Mt. Pleasant. On 5 June 1970 Travis H. McDaniel, then manager at Cape Romain National Wildlife Refuge, made a routine check of nesting birds on Cape Island. As he walked through a Black Skimmer and Gull-billed Tern colony at the south end of the island, he noticed two tern eggs that were appreciably larger than those normally laid by Royal Terns, which are common nesters. During three years at Cape Romain, he had noted that Royal Terns usually lay only one egg. As McDaniel returned to his patrol truck, the birds began to settle back on their eggs. At this time he saw a very large tern dropping down from the air to settle on a nest despite harassing by Black Skimmers. -

Geographical Report of the Crocker Land Expedition, 1913-1917
5.083 (701) Article VL-GEOGRAPHICAL REPORT OF THE CROCKER LAND EXPEDITION, 1913-1917. BY DONALD B. MACMILLAN CONTENTS PAGE INTRODUCTION......................................................... 379 SLEDGE TRIP ON NORTH POLAR SEA, SPRING, 1914 .......................... 384 ASTRONOMICAL OBSERVATIONS-ON NORTH POLAR SEA, 1914 ................ 401 ETAH TO POLAR SEA AND RETURN-MARCH AVERAGES .............. ........ 404 WINTER AND SPRING WORK, 1915-1916 ............. ......................... 404 SPRING WORK OF 1917 .................................... ............ 418 GENERAL SUMMARY ....................................................... 434 INTRODUCTJON The following report embraces the geographical work accomplished by the Crocker Land Expedition during -four years (Summer, 19.13, to Summer, 1917) spent at Etah, NortJaGreenland. Mr. Ekblaw, who was placed in charge of the 1916 expeditin, will present a separate report. The results of the expedition, naturally, depended upon the loca tion of its headquarters. The enforced selection of Etah, North Green- land, seriously handicapped the work of the expedition from start to finish, while the. expenses of the party were more than doubled. The. first accident, the grounding of the Diana upon the coast of Labrador, was a regrettable adventure. The consequent delay, due to unloading, chartering, and reloading, resulted in such a late arrival at Etah that our plans were disarranged. It curtailed in many ways the eageimess of the men to reach their objective point at the head of Flagler Bay, te proposed site of the winter quarters. The leader and his party being but passengers upon a chartered ship was another handicap, since the captain emphatically declared that he would not steam across Smith Sound. There was but one decision to be made, namely: to land upon the North Greenland shore within striking distance of Cape Sabine. -

International Black-Legged Kittiwake Conservation Strategy and Action Plan Acknowledgements Table of Contents
ARCTIC COUNCIL Circumpolar Seabird Expert Group July 2020 International Black-legged Kittiwake Conservation Strategy and Action Plan Acknowledgements Table of Contents Executive Summary ..............................................................................................................................................4 CAFF Designated Agencies: Chapter 1: Introduction .......................................................................................................................................5 • Norwegian Environment Agency, Trondheim, Norway Chapter 2: Ecology of the kittiwake ....................................................................................................................6 • Environment Canada, Ottawa, Canada Species information ...............................................................................................................................................................................................6 • Faroese Museum of Natural History, Tórshavn, Faroe Islands (Kingdom of Denmark) Habitat requirements ............................................................................................................................................................................................6 • Finnish Ministry of the Environment, Helsinki, Finland Life cycle and reproduction ................................................................................................................................................................................7 • Icelandic Institute of Natural -

Holocene Proxy-Climate Data from the Canadian Arctic
Geological Survey of Canada Commission geologique du Canada PAPER 88-22 HOLOCENE PROXY-CLIMATE DATA FROM THE CANADIAN ARCTIC Lynn Ovenden l+I Cana d a 1988 GEOLOGICAL SURVEY OF CANADA PAPER 88-22 HOLOCENE PROXY-CLIMATE DATA FROM THE CANADIAN ARCTIC Lynn Ovenden 1988 Energy, Mines and Energie, Mines et l+I Resources Canada Ressources Canada © Mi ni ster of Supply and Services Canada 1988 Available in Canada through authorized bookstore agent~ and other bookstores or by mail from Canadian Government Publishing Centre Supply and Services Canada Ottawa, Canada KI A OS9 and from Geological Survey of Canada offices: 60 I Booth Street Ottawa. Canada KI A OE8 3303-33rd Street N.W., Calgary, Alberta T2L 2A 7 A deposit copy of thi s publication is also avail able for reference in pu blic libraries across Canada Cat. No. M44-88/ 22E IS BN 0-660-13077-7 Price subject to change wi thout notice Critical reader J. V. Matthews, Jr. Original manuscript submitted - 1988-02- 17 Final version approved for publication - 1988-08-24 CONTENTS Abstract Introduction Background 2 Vascular plant range extensions 2 Arctic peat deposits 2 Eolian deposits 7 Thermokarst and thaw I 0 Conclusions 10 Acknowledgments I 0 References Figure 7 I. An estimate of regional differences in the timing of Holocene peat accumulation on the arctic islands Tables 3 I. Vascular plant range extensions 4 2. Holocene peatbeds 8 3. Organics buried by eolian deposits 9 4. Organics dating thermokarst or ice-wedge collapse HOLOCENE PROXY-CLIMATE DATA FROM THE CANADIAN ARCTIC Abstract A review of diverse reports of the Geological Survey of Canada has produced lists of radiocarbon dated samples from the Canadian Arctic which indicate four climatically significant occurrences during the Holocene: vascular plant range extensions, peat deposits of arctic islands, eolian deposition, and thermokarst and thaw. -

Strategic Environmental Assessment for the Orphan Basin
Orphan Basin Strategic Environmental Assessment Prepared by Prepared for Canada-Newfoundland Offshore Petroleum Board Fifth Floor, TD Place 140 Water Street St. John’s, NL A1C 6H6 11 November 2003 SA767 Orphan Basin Strategic Environmental Assessment Prepared by Prepared for Canada-Newfoundland Offshore Petroleum Board Fifth Floor, TD Place 140 Water Street St. John’s, NL A1C 6H6 11 November 2003 SA767 Orphan Basin Strategic Environmental Assessment Prepared by LGL Limited environmental research associates 388 Kenmount Road St. John’s, NL A1B 4A5 709 754-1992 (p) 709 754-7718 (f) Prepared for Canada-Newfoundland Offshore Petroleum Board Fifth Floor, TD Place 140 Water Street St. John’s, NL A1C 6H6 11 November 2003 SA767 Table of Contents Page Table of Contents........................................................................................................................................ ii List of Tables ............................................................................................................................................. xi List of Figures........................................................................................................................................... xii 1.0 Introduction..................................................................................................................................... 1 2.0 Oil and Gas Activities in Orphan Basin.......................................................................................... 3 2.1 Rights Issuance Process and Call for Bids......................................................................... -

Biodiversity of Michigan's Great Lakes Islands
FILE COPY DO NOT REMOVE Biodiversity of Michigan’s Great Lakes Islands Knowledge, Threats and Protection Judith D. Soule Conservation Research Biologist April 5, 1993 Report for: Land and Water Management Division (CZM Contract 14C-309-3) Prepared by: Michigan Natural Features Inventory Stevens T. Mason Building P.O. Box 30028 Lansing, MI 48909 (517) 3734552 1993-10 F A report of the Michigan Department of Natural Resources pursuant to National Oceanic and Atmospheric Administration Award No. 309-3 BIODWERSITY OF MICHIGAN’S GREAT LAKES ISLANDS Knowledge, Threats and Protection by Judith D. Soule Conservation Research Biologist Prepared by Michigan Natural Features Inventory Fifth floor, Mason Building P.O. Box 30023 Lansing, Michigan 48909 April 5, 1993 for Michigan Department of Natural Resources Land and Water Management Division Coastal Zone Management Program Contract # 14C-309-3 CL] = CD C] t2 CL] C] CL] CD = C = CZJ C] C] C] C] C] C] .TABLE Of CONThNTS TABLE OF CONTENTS I EXECUTIVE SUMMARY iii INTRODUCTION 1 HISTORY AND PHYSICAL RESOURCES 4 Geology and post-glacial history 4 Size, isolation, and climate 6 Human history 7 BIODWERSITY OF THE ISLANDS 8 Rare animals 8 Waterfowl values 8 Other birds and fish 9 Unique plants 10 Shoreline natural communities 10 Threatened, endangered, and exemplary natural features 10 OVERVIEW OF RESEARCH ON MICHIGAN’S GREAT LAKES ISLANDS 13 Island research values 13 Examples of biological research on islands 13 Moose 13 Wolves 14 Deer 14 Colonial nesting waterbirds 14 Island biogeography studies 15 Predator-prey -

Site Fidelity and Colony Dynaimcs of Caspian Terns Nesting at East
AN ABSTRACT OF THE THESIS OF Stefanie Collar for the degree of Master of Science in Wildlife Science presented on December 12, 2013. Title: Site Fidelity and Colony Dynamics of Caspian Terns Nesting at East Sand Island, Columbia River Estuary, Oregon, USA. Abstract approved: ____________________________________________________________________ Daniel D. Roby Fidelity to breeding sites in colonial birds is an adaptive trait thought to have evolved to enhance reproductive success by reducing search time for breeding habitat, allowing earlier nest initiation, facilitating mate retention, and reducing uncertainty of predator presence and food availability. Studying a seabird that has evolved relatively low colony fidelity, such as the Caspian tern, allowed me to explore the influence of stable nesting habitat on fidelity and nest site selection. The Caspian tern (Hydroprogne caspia) breeding colony on East Sand Island (ESI) in the Columbia River estuary is the largest colony of its kind in the world. This colony has experienced a decade of declining nesting success, culminating with the failure of the colony to produce any young in 2011. The objective of my study was to understand the dynamics of this Caspian tern super-colony by investigating the actions of breeding individuals over two seasons, as well as the behavior of the colony as a whole from 2001-2011. I was interested in (1) the degree of nest site fidelity exhibited by breeding terns in successive years and its relationship to reproductive success, and (2) how the interaction of top-down and bottom-up forces influenced average nesting success across the entire colony, and caused the observed trends in nesting success at the East Sand Island colony from 2001 to 2011. -

SHOREBIRDS (Charadriiformes*) CARE MANUAL *Does Not Include Alcidae
SHOREBIRDS (Charadriiformes*) CARE MANUAL *Does not include Alcidae CREATED BY AZA CHARADRIIFORMES TAXON ADVISORY GROUP IN ASSOCIATION WITH AZA ANIMAL WELFARE COMMITTEE Shorebirds (Charadriiformes) Care Manual Shorebirds (Charadriiformes) Care Manual Published by the Association of Zoos and Aquariums in association with the AZA Animal Welfare Committee Formal Citation: AZA Charadriiformes Taxon Advisory Group. (2014). Shorebirds (Charadriiformes) Care Manual. Silver Spring, MD: Association of Zoos and Aquariums. Original Completion Date: October 2013 Authors and Significant Contributors: Aimee Greenebaum: AZA Charadriiformes TAG Vice Chair, Monterey Bay Aquarium, USA Alex Waier: Milwaukee County Zoo, USA Carol Hendrickson: Birmingham Zoo, USA Cindy Pinger: AZA Charadriiformes TAG Chair, Birmingham Zoo, USA CJ McCarty: Oregon Coast Aquarium, USA Heidi Cline: Alaska SeaLife Center, USA Jamie Ries: Central Park Zoo, USA Joe Barkowski: Sedgwick County Zoo, USA Kim Wanders: Monterey Bay Aquarium, USA Mary Carlson: Charadriiformes Program Advisor, Seattle Aquarium, USA Sara Perry: Seattle Aquarium, USA Sara Crook-Martin: Buttonwood Park Zoo, USA Shana R. Lavin, Ph.D.,Wildlife Nutrition Fellow University of Florida, Dept. of Animal Sciences , Walt Disney World Animal Programs Dr. Stephanie McCain: AZA Charadriiformes TAG Veterinarian Advisor, DVM, Birmingham Zoo, USA Phil King: Assiniboine Park Zoo, Canada Reviewers: Dr. Mike Murray (Monterey Bay Aquarium, USA) John C. Anderson (Seattle Aquarium volunteer) Kristina Neuman (Point Blue Conservation Science) Sarah Saunders (Conservation Biology Graduate Program,University of Minnesota) AZA Staff Editors: Maya Seaman, MS, Animal Care Manual Editing Consultant Candice Dorsey, PhD, Director of Animal Programs Debborah Luke, PhD, Vice President, Conservation & Science Cover Photo Credits: Jeff Pribble Disclaimer: This manual presents a compilation of knowledge provided by recognized animal experts based on the current science, practice, and technology of animal management. -

Pacific Seabirds
PACIFIC SEABIRDS A Publication of the Pacific Seabird Group Volume 36 Number 2 Fall 2009 PACIFIC SEABIRD GROUP Dedicated to the Study and Conservation of Pacific Seabirds and Their Environment The Pacific Seabird Group (PSG) was formed in 1972 due to the need for better communication among Pacific seabird researchers. PSG provides a forum for the research activities of its members, promotes the conservation of seabirds, and informs members and the public of issues relating to Pacific Ocean seabirds and their environment. PSG members include research scientists, conservation professionals, and members of the public from all parts of the Pacific Ocean. The group also welcomes seabird professionals and enthusiasts in other parts of the world. PSG holds annual meetings at which scientific papers and symposia are presented; abstracts for meetings are published on our web site. The group is active in promoting conservation of seabirds, include seabird/fisheries interactions, monitoring of seabird populations, seabird restoration following oil spills, establishment of seabird sanctuaries, and endangered species. Policy statements are issued on conservation issues of critical importance. PSG’s journals are Pacific Seabirds (formerly the PSG Bulletin) and Marine Ornithology. Other publications include symposium volumes and technical reports; these are listed near the back of this issue. PSG is a member of the International Union for Conservation of Nature (IUCN), the Ornithological Council, and the American Bird Conservancy. Annual dues for membership are $30 (individual and family); $24 (student, undergraduate and graduate); and $900 (Life Membership, payable in five $180 installments). Dues are payable to the Treasurer; see the PSG web site, or the Membership Order Form next to inside back cover. -

Hydroprogne Caspia
Prepared in cooperation with the U.S. Army Corps of Engineers and the Bureau of Reclamation Evaluation of Caspian Tern (Hydroprogne caspia) and Snowy Plover (Charadrius alexandrinus nivosus) Nesting on Modified Islands at the Don Edwards San Francisco Bay National Wildlife Refuge, California—2016 Annual Report Open-File Report 2017–1055 U.S. Department of the Interior U.S. Geological Survey Cover: Photograph showing Caspian tern (Hydroprogne caspia) chicks and decoy on an enhanced island in Pond SF2 of the Don Edwards San Francisco Bay National Wildlife Refuge, California. Photograph by Crystal Shore, U.S. Geological Survey, May 11, 2016. Evaluation of Caspian Tern (Hydroprogne caspia) and Snowy Plover (Charadrius alexandrinus nivosus) Nesting on Modified Islands at the Don Edwards San Francisco Bay National Wildlife Refuge, California— 2016 Annual Report By C. Alex Hartman, Joshua T. Ackerman, Mark P. Herzog, Cheryl Strong, David Trachtenbarg, and Crystal A. Shore Prepared in cooperation with the U.S. Army Corps of Engineers and the Bureau of Reclamation Open-File Report 2017–1055 U.S. Department of the Interior U.S. Geological Survey U.S. Department of the Interior RYAN K. ZINKE, Secretary U.S. Geological Survey William H. Werkheiser, Acting Director U.S. Geological Survey, Reston, Virginia: 2017 For more information on the USGS—the Federal source for science about the Earth, its natural and living resources, natural hazards, and the environment—visit https://www.usgs.gov/ or call 1–888–ASK–USGS (1–888–275–8747). For an overview of USGS information products, including maps, imagery, and publications, visit https:/store.usgs.gov.