Ache Ttn Ache Nfkb1 Il6 Nfkb1 Ca2 Tnf Ca2 Ca7 Ace Maoa
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Activated Peripheral-Blood-Derived Mononuclear Cells
Transcription factor expression in lipopolysaccharide- activated peripheral-blood-derived mononuclear cells Jared C. Roach*†, Kelly D. Smith*‡, Katie L. Strobe*, Stephanie M. Nissen*, Christian D. Haudenschild§, Daixing Zhou§, Thomas J. Vasicek¶, G. A. Heldʈ, Gustavo A. Stolovitzkyʈ, Leroy E. Hood*†, and Alan Aderem* *Institute for Systems Biology, 1441 North 34th Street, Seattle, WA 98103; ‡Department of Pathology, University of Washington, Seattle, WA 98195; §Illumina, 25861 Industrial Boulevard, Hayward, CA 94545; ¶Medtronic, 710 Medtronic Parkway, Minneapolis, MN 55432; and ʈIBM Computational Biology Center, P.O. Box 218, Yorktown Heights, NY 10598 Contributed by Leroy E. Hood, August 21, 2007 (sent for review January 7, 2007) Transcription factors play a key role in integrating and modulating system. In this model system, we activated peripheral-blood-derived biological information. In this study, we comprehensively measured mononuclear cells, which can be loosely termed ‘‘macrophages,’’ the changing abundances of mRNAs over a time course of activation with lipopolysaccharide (LPS). We focused on the precise mea- of human peripheral-blood-derived mononuclear cells (‘‘macro- surement of mRNA concentrations. There is currently no high- phages’’) with lipopolysaccharide. Global and dynamic analysis of throughput technology that can precisely and sensitively measure all transcription factors in response to a physiological stimulus has yet to mRNAs in a system, although such technologies are likely to be be achieved in a human system, and our efforts significantly available in the near future. To demonstrate the potential utility of advanced this goal. We used multiple global high-throughput tech- such technologies, and to motivate their development and encour- nologies for measuring mRNA levels, including massively parallel age their use, we produced data from a combination of two distinct signature sequencing and GeneChip microarrays. -

Cyclin D1 Is a Direct Transcriptional Target of GATA3 in Neuroblastoma Tumor Cells
Oncogene (2010) 29, 2739–2745 & 2010 Macmillan Publishers Limited All rights reserved 0950-9232/10 $32.00 www.nature.com/onc SHORT COMMUNICATION Cyclin D1 is a direct transcriptional target of GATA3 in neuroblastoma tumor cells JJ Molenaar1,2, ME Ebus1, J Koster1, E Santo1, D Geerts1, R Versteeg1 and HN Caron2 1Department of Human Genetics, Academic Medical Center, University of Amsterdam, Amsterdam, The Netherlands and 2Department of Pediatric Oncology, Emma Kinderziekenhuis, Academic Medical Center, University of Amsterdam, Amsterdam, The Netherlands Almost all neuroblastoma tumors express excess levels of 2000). Several checkpoints normally prevent premature Cyclin D1 (CCND1) compared to normal tissues and cell-cycle progression and cell division. The crucial G1 other tumor types. Only a small percentage of these entry point is controlled by the D-type Cyclins that can neuroblastoma tumors have high-level amplification of the activate CDK4/6 that in turn phosphorylate the pRb Cyclin D1 gene. The other neuroblastoma tumors have protein. This results in a release of the E2F transcription equally high Cyclin D1 expression without amplification. factor that causes transcriptional upregulation of Silencing of Cyclin D1 expression was previously found to numerous genes involved in further progression of the trigger differentiation of neuroblastoma cells. Over- cell cycle (Sherr, 1996). expression of Cyclin D1 is therefore one of the most Neuroblastomas are embryonal tumors that originate frequent mechanisms with a postulated function in neuro- from precursor cells of the sympathetic nervous system. blastoma pathogenesis. The cause for the Cyclin D1 This tumor has a very poor prognosis and despite the overexpression is unknown. -

Ten Commandments for a Good Scientist
Unravelling the mechanism of differential biological responses induced by food-borne xeno- and phyto-estrogenic compounds Ana María Sotoca Covaleda Wageningen 2010 Thesis committee Thesis supervisors Prof. dr. ir. Ivonne M.C.M. Rietjens Professor of Toxicology Wageningen University Prof. dr. Albertinka J. Murk Personal chair at the sub-department of Toxicology Wageningen University Thesis co-supervisor Dr. ir. Jacques J.M. Vervoort Associate professor at the Laboratory of Biochemistry Wageningen University Other members Prof. dr. Michael R. Muller, Wageningen University Prof. dr. ir. Huub F.J. Savelkoul, Wageningen University Prof. dr. Everardus J. van Zoelen, Radboud University Nijmegen Dr. ir. Toine F.H. Bovee, RIKILT, Wageningen This research was conducted under the auspices of the Graduate School VLAG Unravelling the mechanism of differential biological responses induced by food-borne xeno- and phyto-estrogenic compounds Ana María Sotoca Covaleda Thesis submitted in fulfillment of the requirements for the degree of doctor at Wageningen University by the authority of the Rector Magnificus Prof. dr. M.J. Kropff, in the presence of the Thesis Committee appointed by the Academic Board to be defended in public on Tuesday 14 September 2010 at 4 p.m. in the Aula Unravelling the mechanism of differential biological responses induced by food-borne xeno- and phyto-estrogenic compounds. Ana María Sotoca Covaleda Thesis Wageningen University, Wageningen, The Netherlands, 2010, With references, and with summary in Dutch. ISBN: 978-90-8585-707-5 “Caminante no hay camino, se hace camino al andar. Al andar se hace camino, y al volver la vista atrás se ve la senda que nunca se ha de volver a pisar” - Antonio Machado – A mi madre. -

Functional Genomics Analysis of Vitamin D Effects on CD4+ T Cells In
Functional genomics analysis of vitamin D effects PNAS PLUS on CD4+ T cells in vivo in experimental autoimmune encephalomyelitis Manuel Zeitelhofera,b, Milena Z. Adzemovica, David Gomez-Cabreroc,d,e, Petra Bergmana, Sonja Hochmeisterf, Marie N’diayea, Atul Paulsona, Sabrina Ruhrmanna, Malin Almgrena, Jesper N. Tegnérc,d,g, Tomas J. Ekströma, André Ortlieb Guerreiro-Cacaisa, and Maja Jagodica,1 aDepartment of Clinical Neuroscience, Center for Molecular Medicine, Karolinska Institutet, 171 76 Stockholm, Sweden; bVascular Biology Unit, Department of Medical Biochemistry and Biophysics, Karolinska Institutet, 171 77 Stockholm, Sweden; cUnit of Computational Medicine, Department of Medicine, Solna, Center for Molecular Medicine, Karolinska Institutet, 171 76 Stockholm, Sweden; dScience for Life Laboratory, 171 21 Solna, Sweden; eMucosal and Salivary Biology Division, King’s College London Dental Institute, London SE1 9RT, United Kingdom; fDepartment of General Neurology, Medical University of Graz, 8036 Graz, Austria; and gBiological and Environmental Sciences and Engineering Division, Computer, Electrical and Mathematical Sciences and Engineering Division, King Abdullah University of Science and Technology, 23955 Thuwal, Kingdom of Saudi Arabia Edited by Tomas G. M. Hokfelt, Karolinska Institutet, Stockholm, Sweden, and approved January 19, 2017 (received for review September 24, 2016) Vitamin D exerts multiple immunomodulatory functions and has autoimmune destruction of myelin, axonal loss, and brain atro- been implicated in the etiology and treatment of several autoim- phy (6). Increased risk of developing MS has been described in mune diseases, including multiple sclerosis (MS). We have previously carriers of rare and common variants of the CYP27B gene (7, 8), reported that in juvenile/adolescent rats, vitamin D supplementation which encodes the enzyme that catalyzes the last step in con- protects from experimental autoimmune encephalomyelitis (EAE), a verting vitamin D to its active form, from 25(OH)D3 to 1,25 model of MS. -

Mirc11 Disrupts Inflammatory but Not Cytotoxic Responses of NK Cells
Published OnlineFirst September 12, 2019; DOI: 10.1158/2326-6066.CIR-18-0934 Research Article Cancer Immunology Research Mirc11 Disrupts Inflammatory but Not Cytotoxic Responses of NK Cells Arash Nanbakhsh1, Anupallavi Srinivasamani1, Sandra Holzhauer2, Matthew J. Riese2,3,4, Yongwei Zheng5, Demin Wang4,5, Robert Burns6, Michael H. Reimer7,8, Sridhar Rao7,8, Angela Lemke9,10, Shirng-Wern Tsaih9,10, Michael J. Flister9,10, Shunhua Lao1,11, Richard Dahl12, Monica S. Thakar1,11, and Subramaniam Malarkannan1,3,4,9,11 Abstract Natural killer (NK) cells generate proinflammatory cyto- g–dependent clearance of Listeria monocytogenes or B16F10 kines that are required to contain infections and tumor melanoma in vivo by NK cells. These functional changes growth. However, the posttranscriptional mechanisms that resulted from Mirc11 silencing ubiquitin modifiers A20, regulate NK cell functions are not fully understood. Here, we Cbl-b, and Itch, allowing TRAF6-dependent activation of define the role of the microRNA cluster known as Mirc11 NF-kB and AP-1. Lack of Mirc11 caused increased translation (which includes miRNA-23a, miRNA-24a, and miRNA-27a) of A20, Cbl-b, and Itch proteins, resulting in deubiquityla- in NK cell–mediated proinflammatory responses. Absence tion of scaffolding K63 and addition of degradative K48 of Mirc11 did not alter the development or the antitumor moieties on TRAF6. Collectively, our results describe a func- cytotoxicity of NK cells. However, loss of Mirc11 reduced tion of Mirc11 that regulates generation of proinflammatory generation of proinflammatory factors in vitro and interferon- cytokines from effector lymphocytes. Introduction TRAF2 and TRAF6 promote K63-linked polyubiquitination that is required for subcellular localization of the substrates (20), Natural killer (NK) cells generate proinflammatory factors and and subsequent activation of NF-kB (21) and AP-1 (22). -

Original Article a Database and Functional Annotation of NF-Κb Target Genes
Int J Clin Exp Med 2016;9(5):7986-7995 www.ijcem.com /ISSN:1940-5901/IJCEM0019172 Original Article A database and functional annotation of NF-κB target genes Yang Yang, Jian Wu, Jinke Wang The State Key Laboratory of Bioelectronics, Southeast University, Nanjing 210096, People’s Republic of China Received November 4, 2015; Accepted February 10, 2016; Epub May 15, 2016; Published May 30, 2016 Abstract: Backgrounds: The previous studies show that the transcription factor NF-κB always be induced by many inducers, and can regulate the expressions of many genes. The aim of the present study is to explore the database and functional annotation of NF-κB target genes. Methods: In this study, we manually collected the most complete listing of all NF-κB target genes identified to date, including the NF-κB microRNA target genes and built the database of NF-κB target genes with the detailed information of each target gene and annotated it by DAVID tools. Results: The NF-κB target genes database was established (http://tfdb.seu.edu.cn/nfkb/). The collected data confirmed that NF-κB maintains multitudinous biological functions and possesses the considerable complexity and diversity in regulation the expression of corresponding target genes set. The data showed that the NF-κB was a central regula- tor of the stress response, immune response and cellular metabolic processes. NF-κB involved in bone disease, immunological disease and cardiovascular disease, various cancers and nervous disease. NF-κB can modulate the expression activity of other transcriptional factors. Inhibition of IKK and IκBα phosphorylation, the decrease of nuclear translocation of p65 and the reduction of intracellular glutathione level determined the up-regulation or down-regulation of expression of NF-κB target genes. -

Pathway and Network Analysis of Somatic Mutations Across Cancer
Network Analysis of Mutaons Across Cancer Types Ben Raphael Fabio Vandin, Max Leiserson, Hsin-Ta Wu Department of Computer Science Center for Computaonal Molecular Biology Significantly Mutated Genes Muta#on Matrix Stascal test Genes Paents Frequency Number Paents Study Num. Samples Num. SMG TCGA Ovarian (2011) 316 10 TCGA Breast (2012) 510 35 TCGA Colorectal (2012) 276 32 background mutaon rate (BMR), gene specific effects, etc. Significantly Mutated Genes à Pathways Stascal test Frequency Number Paents TCGA Colorectal (Nature 2012) TCGA Ovarian (Nature 2011) background mutaon rate (BMR), gene specific effects, etc. Advantages of Large Datasets Prior knowledge of groups of genes Genes Paents Known pathways Interac3on Network None Prior knowledge • Novel pathways or interac3ons between pathways (crosstalk) • Topology of interac3ons Two Algorithms Prior knowledge of groups of genes Genes Paents Known pathways Interac3on Network None Prior knowledge Number of Hypotheses HotNet subnetworks of Dendrix interac3on network Exclusive gene sets HotNet: Problem Defini3on Given: 1. Network G = (V, E) V = genes. E = interac3ons b/w genes 2. Binary mutaon matrix Genes = mutated = not mutated Paents Find: Connected subnetworks mutated in a significant number of paents. Subnetwork Properes Mutaon frequency/score AND network topology Frequency Number Paents • Moderate frequency/score • High frequency/score • Highly connected • Connected through high-degree node. Example: TP53 has 238 neighbors in HPRD network Mutated subnetworks: HotNet* Muta#on Matrix Human Interac#on Network Genes = mutated genes Paents (1) Muta#on à heat diffusion Extract “significantly hot” subnetworks Hot (2) Cold *F. Vandin, E. Upfal, and B. J. Raphael. J. Comp.Biol. (2011). Also RECOMB (2010). Stas3cal Test Muta#on Matrix Random Binary Matrix Genes Genes Paents Paents Xs = number of subnetworks ≥ s genes Two-stage mul-hypothesis test: Rigorously bound FDR. -

Supplementary Table 1
Supplementary Table 1. 492 genes are unique to 0 h post-heat timepoint. The name, p-value, fold change, location and family of each gene are indicated. Genes were filtered for an absolute value log2 ration 1.5 and a significance value of p ≤ 0.05. Symbol p-value Log Gene Name Location Family Ratio ABCA13 1.87E-02 3.292 ATP-binding cassette, sub-family unknown transporter A (ABC1), member 13 ABCB1 1.93E-02 −1.819 ATP-binding cassette, sub-family Plasma transporter B (MDR/TAP), member 1 Membrane ABCC3 2.83E-02 2.016 ATP-binding cassette, sub-family Plasma transporter C (CFTR/MRP), member 3 Membrane ABHD6 7.79E-03 −2.717 abhydrolase domain containing 6 Cytoplasm enzyme ACAT1 4.10E-02 3.009 acetyl-CoA acetyltransferase 1 Cytoplasm enzyme ACBD4 2.66E-03 1.722 acyl-CoA binding domain unknown other containing 4 ACSL5 1.86E-02 −2.876 acyl-CoA synthetase long-chain Cytoplasm enzyme family member 5 ADAM23 3.33E-02 −3.008 ADAM metallopeptidase domain Plasma peptidase 23 Membrane ADAM29 5.58E-03 3.463 ADAM metallopeptidase domain Plasma peptidase 29 Membrane ADAMTS17 2.67E-04 3.051 ADAM metallopeptidase with Extracellular other thrombospondin type 1 motif, 17 Space ADCYAP1R1 1.20E-02 1.848 adenylate cyclase activating Plasma G-protein polypeptide 1 (pituitary) receptor Membrane coupled type I receptor ADH6 (includes 4.02E-02 −1.845 alcohol dehydrogenase 6 (class Cytoplasm enzyme EG:130) V) AHSA2 1.54E-04 −1.6 AHA1, activator of heat shock unknown other 90kDa protein ATPase homolog 2 (yeast) AK5 3.32E-02 1.658 adenylate kinase 5 Cytoplasm kinase AK7 -

Effects of Targeting the Transcription Factors Ikaros and Aiolos on B Cell Activation and Differentiation in Systemic Lupus Erythematosus
Immunology and inflammation Lupus Sci Med: first published as 10.1136/lupus-2020-000445 on 16 March 2021. Downloaded from Effects of targeting the transcription factors Ikaros and Aiolos on B cell activation and differentiation in systemic lupus erythematosus Felice Rivellese ,1 Sotiria Manou- Stathopoulou,1 Daniele Mauro,1 Katriona Goldmann,1 Debasish Pyne,2 Ravindra Rajakariar,3 Patrick Gordon,4 Peter Schafer,5 Michele Bombardieri,1 Costantino Pitzalis,1 Myles J Lewis 1 To cite: Rivellese F, ABSTRACT Manou- Stathopoulou S, Objective To evaluate the effects of targeting Ikaros and Key messages Mauro D, et al. Effects of Aiolos by cereblon modulator iberdomide on the activation What is already known about this subject? targeting the transcription and differentiation of B- cells from patients with systemic factors Ikaros and Aiolos The transcription factors Ikaros and Aiolos, which lupus erythematosus (SLE). ► on B cell activation and are critical for B cell differentiation, are implicated in Methods CD19+ B- cells isolated from the peripheral differentiation in systemic systemic lupus erythematosus (SLE) pathogenesis. blood of patients with SLE (n=41) were cultured with lupus erythematosus. Targeting Ikaros and Aiolos using the cereblon mod- TLR7 ligand resiquimod ±IFNα together with iberdomide ► Lupus Science & Medicine ulator iberdomide has been proposed as a promising 2021;8:e000445. doi:10.1136/ or control from day 0 (n=16). Additionally, in vitro B- cell therapeutic agent. lupus-2020-000445 differentiation was induced by stimulation with IL-2/IL-10/ IL-15/CD40L/resiquimod with iberdomide or control, given What does this study add? at day 0 or at day 4. -

NFKB1 and Cancer: Friend Or Foe?
cells Review NFKB1 and Cancer: Friend or Foe? Julia Concetti and Caroline L. Wilson * Newcastle Fibrosis Research Group, Institute of Cellular Medicine, Newcastle University, Newcastle upon Tyne, Tyne and Wear NE2 4HH, UK; [email protected] * Correspondence: [email protected]; Tel.: +44-191-208-8590 Received: 15 August 2018; Accepted: 4 September 2018; Published: 7 September 2018 Abstract: Current evidence strongly suggests that aberrant activation of the NF-κB signalling pathway is associated with carcinogenesis. A number of key cellular processes are governed by the effectors of this pathway, including immune responses and apoptosis, both crucial in the development of cancer. Therefore, it is not surprising that dysregulated and chronic NF-κB signalling can have a profound impact on cellular homeostasis. Here we discuss NFKB1 (p105/p50), one of the five subunits of NF-κB, widely implicated in carcinogenesis, in some cases driving cancer progression and in others acting as a tumour-suppressor. The complexity of the role of this subunit lies in the multiple dimeric combination possibilities as well as the different interacting co-factors, which dictate whether gene transcription is activated or repressed, in a cell and organ-specific manner. This review highlights the multiple roles of NFKB1 in the development and progression of different cancers, and the considerations to make when attempting to manipulate NF-κB as a potential cancer therapy. Keywords: NF-κB; NFKB1; p105/p50; Bcl-3; cancer; inflammation; apoptosis 1. Introduction One of the emerging questions in cancer biology is: “How are inflammation and dysregulated immune responses linked to cancer?” It is now widely accepted that chronic inflammation and infection represent major risk factors for certain cancers. -

Identi Cation of Phenotype-Speci C Networks from Paired Gene Expression-Cell Shape Imaging Data
bioRxiv preprint doi: https://doi.org/10.1101/2021.02.11.430597; this version posted July 17, 2021. The copyright holder for this preprint (which was not certified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available under aCC-BY 4.0 International license. Identification of phenotype-specific networks from paired gene expression-cell shape imaging data Charlie George Barker1, Eirini Petsalaki1, Girolamo Giudice1, Julia Sero2, Emmanuel Nsa Ekpenyong1, Chris Bakal3, Evangelia Petsalaki1∗ 1European Molecular Biology Laboratory-European Bioinformatics Institute, Hinxton CB10 1SD, UK 2University of Bath, Claverton Down, Bath BA2 7AY, UK 3Institute of Cancer Research, 237 Fulham Road, London, SW3 6JB, UK ∗Lead Contact/Correspondence: Evangelia Petsalaki, [email protected] Abstract The morphology of breast cancer cells is often used as an indicator of tumour severity and prognosis. Additionally, morphology can be used to identify more fine-grained, molecular developments within a cancer cell, such as transcriptomic changes and signaling pathway activity. Delineating the interface be- tween morphology and signaling is important to understand the mechanical cues that a cell processes in order to undergo epithelial-to-mesenchymal transition and consequently metastasize. However, the exact regulatory systems that define these changes remain poorly characterised. In this study, we em- ploy a network-systems approach to integrate imaging data and RNA-seq expression data. Our workflow allows the discovery of unbiased and context-specific gene expression signatures and cell signaling sub- networks relevant to the regulation of cell shape, rather than focusing on the identification of previously known, but not always representative, pathways. -
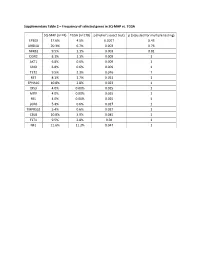
Supplementary Table 2 – Frequency of Selected Genes in SQ-MAP Vs
Supplementary Table 2 – Frequency of selected genes in SQ-MAP vs. TCGA SQ-MAP (n=74) TCGA (n=178) p (Fisher’s exact test) p (adjusted for multiple testing) EP300 17.6% 4.5% 0.0017 0.43 ARID1A 20.3% 6.7% 0.003 0.76 NFKB1 9.5% 1.1% 0.003 0.81 DDR2 8.1% 1.1% 0.009 1 AKT1 6.8% 0.6% 0.009 1 SMO 6.8% 0.6% 0.009 1 TET2 9.5% 2.3% 0.016 1 RET 8.1% 1.7% 0.021 1 EPHA10 10.8% 2.8% 0.023 1 DIS3 4.0% 0.00% 0.025 1 MITF 4.0% 0.00% 0.025 1 REL 4.0% 0.00% 0.025 1 LGR6 5.4% 0.6% 0.027 1 TMPRSS2 5.4% 0.6% 0.027 1 CBLB 10.8% 3.9% 0.045 1 FLT4 9.5% 2.8% 0.04 1 NF1 21.6% 11.2% 0.047 1 Supplementary Table 3 – Clinical characteristics of patients with resected brain metastases ID Age Sex Race Smoking status Pack-years Treatment cisplatin + docetaxel -> cisplatin + PP1 51 Female White Former 10 gemcitabine -> sequential RT; gemcitabine; nab-paclitaxel PP2 88 Male White Former 50 carboplatin + paclitaxel PP3 62 Female White Current 33 cisplatin + etoposide + concurrent RT PP4 66 Female White Former 20 cisplatin + gemcitabine carboplatin + paclitaxel + concurrent PP5 67 Female White Former 50 RT PP6 77 Male White Current 53 carboplatin + paclitaxel Supplementary Table 4 – Genes exhibiting clonal evolution from primary lung tumors and matched brain metastases.