Effects of Intraspecific Competition on Sheltering Behavior in the Desert Sand Scorpion
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Beck's Desert Scorpion
Molecular Phylogenetics and Evolution 69 (2013) 502–513 Contents lists available at ScienceDirect Molecular Phylogenetics and Evolution journal homepage: www.elsevier.com/locate/ympev Phylogeography of Beck’s Desert Scorpion, Paruroctonus becki, reveals Pliocene diversification in the Eastern California Shear Zone and postglacial expansion in the Great Basin Desert ⇑ Matthew R. Graham a, , Jef R. Jaeger a, Lorenzo Prendini b, Brett R. Riddle a a School of Life Sciences, University of Nevada Las Vegas, 4505 South Maryland Parkway, Las Vegas, NV 89154-4004, USA b Division of Invertebrate Zoology, American Museum of Natural History, Central Park West at 79th Street, New York, NY 10024-5192, USA article info abstract Article history: The distribution of Beck’s Desert Scorpion, Paruroctonus becki (Gertsch and Allred, 1965), spans the Received 12 November 2012 ‘warm’ Mojave Desert and the western portion of the ‘cold’ Great Basin Desert. We used genetic analyses Revised 10 July 2013 and species distribution modeling to test whether P. becki persisted in the Great Basin Desert during the Accepted 29 July 2013 Last Glacial Maximum (LGM), or colonized the area as glacial conditions retreated and the climate Available online 9 August 2013 warmed. Phylogenetic and network analyses of mitochondrial cytochrome c oxidase 1 (cox1), 16S rDNA, and nuclear internal transcribed spacer (ITS-2) DNA sequences uncovered five geographically-structured Keywords: groups in P. becki with varying degrees of statistical support. Molecular clock estimates and the geograph- Biogeography ical arrangement of three of the groups suggested that Pliocene geological events in the tectonically Basin and range COI dynamic Eastern California Shear Zone may have driven diversification by vicariance. -

Chemosensory Physiology and Behavior of the Desert Sand Scorpion
AN ABSTRACT OF THE THESIS OF Douglas Dean Gaffin for the degree of Doctor of Philosophy in Zoology presented on September 23. 1993. Title: Chemosensory Physiology and Behavior of the Desert Sand Scorpion, Paruroctonus mesaensis. Redacted for Privacy Abstract approved: Philip H. Brownell This is a neuroethological study of two major chemosensory systems found in all scorpions - the large, ventral appendages, called pectines, found uniquely in this taxon, and setaform chemoreceptors of the tarsal leg segments. These sensory organs are closely associated with the substrate and their microstructure suggests specialized function in gustation or near- field olfaction of chemical substances on dry surfaces. In this study I present behavioral and electrophysiological evidence that the numerous peg sensilla on the pectines are important chemosensory channels in scorpions and probably fill similar functional roles to antenna' sensilla of rnandibulate arthropods. The subject of these investigations was the desert sand scorpion Paruroctonus mesaensis. Sand scorpions displayed vigorous, stereotyped behavior in response to substrates treated with water and chemicals derived from conspecific scorpions. Ablation studies showed the pectines are important in the detection of substrate-borne pheromonal signals while the tarsal chemosensory hairs are important detectors of substrate water. Electrophysiological investigation of individual peg sensilla on the pectines showed these structures are sensitive to chemostimulants applied directly to the sensillar tip or blown across its pore. Neurons within each peg gave characteristic patterns of response to organic stimuli of various classification (alkanes, alcohols, aldehydes, ketones, esters) that were generally independent of carbon chain length (C2 to C12). Tarsal hair sensilla were responsive to water applied directly to the hair tips. -

A New Species of Paruroctonus from Coastal California (Scorpiones: Vaejovidae)
PAN-PACIFIC ENTOMOLOGIST 63(4), 1987, pp. 329-332 A New Species of Paruroctonus from Coastal California (Scorpiones: Vaejovidae) STANLEY C. WILLIAMS Department of Biology, San Francisco State University, San Francisco, California 94132. Abstract.—A new species of scorpion from coastal California is described and named Paruroctonus maritimus Williams. Its nearest relatives appear to be Paruroctonus boreus (Girard) and Paruroctonus silvestrii (Borelli). A few years ago a small series of Paruroctonus was collected under cardboard and other surface debris by Roy Johnson, along railroad tracks at Sea Side, Monterey County, California. At the time, this sample posed certain problems of interpretation, although it appeared to be Paruroctonus boreus (Girard). Since the collection site was well outside of the known distribution of P. boreus, restricted in area, and ecologically disturbed, the identification of this species was tentative. Reinvestigation of these specimens indicates they are not P. boreus, but an undescribed new species that appears to inhabit the coastal sand dune community. This new species is here described and named. The measurements cited are as defined by Williams (1980). Paruroctonus maritimus Williams, NEW SPECIES (Fig. 1, Table 1) Diagnosis.—Members of subgenus Paruroctonus. Total length up to 50 mm. base color of exoskeleton pale yellow with contrasting dusky-black marbling dorsally on carapace and mesosoma, dark marbling not extending to posterior margin of mesosomal terga, metasoma with ventral and ventrolateral keels outlined in dusky pigment; frontal margin of carapace convex; pectine teeth 24-27 in males, 18-20 in females; metasoma with ventral keels smooth to obsolete on I, smooth on II—IV, serrate on V; metasoma with ventrolateral keels smooth to granular on I—II, smooth on III, smooth to crenulate on IV; chela with supernumerary denticles 6 on fixed, 7 on movable finger; chela with primary row denticles divided into 6 subrows on fixed finger, 7 subrows on movable finger. -
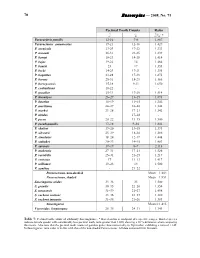
Euscorpius — 2008, No. 71 70
70 Euscorpius — 2008, No. 71 Pectinal Tooth Counts Ratio ♂ ♀ ♂/♀ * Paravaejovis pumilis 12-16 7-8 1.867 Paruroctonus ammonastes 19-21 12-16 1.429 P. arenicola 23-29 17-22 1.333 P. arnaudi 26-31 21-25 1.239 P. baergi 18-23 14-15 1.414 P. bajae 19-22 14 1.464 P. bantai 23 17 1.353 P. becki 24-29 17-21 1.395 P. boquillas 23-28 17-23 1.275 P. boreus 25-31 18-23 1.366 P. borregoensis 15-18 9-11 1.650 P. coahuilanus 18-22 - - P. gracilior 25-31 17-20 1.514 P. hirsutipes 26–27 13–15 1.893 P. luteolus 16–19 11–15 1.346 P. maritimus 24–27 18–20 1.342 P. marksi 23–28 17–21 1.342 P. nitidus - 17–18 - P. pecos 20–22 13–15 1.500 P. pseudopumilis 17–18 9–10 1.842 P. shulovi 19–20 13–15 1.393 P. silvestrii 25–29 18-24 1.286 P. simulatus 18–24 12-17 1.448 P. stahnkei 20–23 14–15 1.483 P. surensis 17–19 8–9 2.118 P. utahensis 27–31 17–21 1.526 P. variabilis 25–31 21–25 1.217 P. ventosus 17 11–13 1.417 P. williamsi 19–23 14 1.500 P. xanthus - 21–22 - Paruroctonus, non-shaded Mean = 1.402 Paruroctonus, shaded Mean = 1.951 Smeringurus aridus 33–36 23 1.500 S. grandis 30–35 22–26 1.354 S. mesaensis 31–39 21–27 1.458 S. -

UG ETD Template
Exploring genome size diversity in arachnid taxa by Haley Lauren Yorke A Thesis presented to The University of Guelph In partial fulfilment of requirements for the degree of Master of Science in Integrative Biology Guelph, Ontario, Canada © Haley L. Yorke, January, 2020 ABSTRACT EXPLORING GENOME SIZE DIVERSITY IN ARACHNID TAXA Haley L. Yorke Advisor(s): University of Guelph, 2019 Dr. T. Ryan Gregory This study investigates the general ranges and patterns of arachnid genome size diversity within and among major arachnid groups, with a focus on tarantulas (Theraphosidae) and scorpions (Scorpiones), and an examination of how genome size relates to body size, growth rate, longevity, and geographical latitude. I produced genome size measurements for 151 new species, including 108 tarantulas (Theraphosidae), 20 scorpions (Scorpiones), 17 whip-spiders (Amblypygi), 1 vinegaroon (Thelyphonida), and 5 non-arachnid relatives – centipedes (Chilopoda). I also developed a new methodology for non-lethal sampling of arachnid species that is inexpensive and portable, which will hopefully improve access to live specimens in zoological or hobbyist collections. I found that arachnid genome size is positively correlated with longevity, body size and growth rate. Despite this general trend, genome size was found to negatively correlate with longevity and growth rate in tarantulas (Theraphosidae). Tarantula (Theraphosidae) genome size was correlated negatively with latitude. iii ACKNOWLEDGEMENTS This study would not have been possible without access to specimens from the collections of both Amanda Gollaway and Martin Gamache of Tarantula Canada, and also of Patrik de los Reyes – thank you for trusting me with your valuable and much- loved animals, for welcoming me into your homes and places of business, and for your expert opinions on arachnid husbandry and biology. -

The Effects of Magnetic Fields on the Desert Grassland Scorpion, Paruroctonus Utahensis
Magnetoreception in scorpions; pg 1 of 5 The effects of magnetic fields on the desert grassland scorpion, Paruroctonus utahensis. A biology cornerstone manuscript by: Quincey Ward, Robert Kunkel, Christopher Heigle Spring 2016 The Department of Biology; University of Oklahoma; Norman, Oklahoma 73019 Course instructors: Doug Gaffin, Brad Brayfield Summary Recent studies have shown that desert ants, Cataglyphis noda, use magnetic fields as directional information to return to their nests. This process, known as magnetoreception, is speculated to be the result of UV-A/blue light photoreceptors called cryptochromes inducing radical pair reactions in the retinas when exposed to light under 420 nm. Although research has been conducted on magnetoreception in the arthropod subphylum Hexapoda, the Chelicerata subphylum had yet to be studied. In this study, we tested for traces of magnetoreception in fifteen female dessert sand scorpions (Paruroctonus uthahensis). The scorpions were placed in a segmented arena with a treated zone situated between two Helmholtz coils. We tested the amount of time each scorpion spent and their velocities in each zone when exposed to three magnetic fields: 0, 100, and 500 µT, . We predicted there would be a change in animal behavior in the treated zone when a magnetic field was present compared to the control trials. Our results showed no evidence of the animals occupying the treated zone more frequently than the untreated zone. Under the conditions of these experiments, scorpions did not show signs of magnetoreception. For future tests on scorpion navigation, we suggest researching e-vectors, which are used by bees as an internal compass. Introduction proteins known as cryptochromes to initiate the radical- pair reactions necessary to induce the ability to use While we consider ourselves to be an advanced species, magnetic fields as a way of perceiving direction and humans cannot detect changes in Earth’s magnetic fields. -

Phylogeography and Geographical Variation of Behavioral
PHYLOGEOGRAPHY AND GEOGRAPHICAL VARIATION OF BEHAVIORAL AND MORPHOLOGICAL CHARACTERISTICS IN PARUROCTONUS BOREUS by ABRAHAM MILLER Presented to the Faculty of the Graduate School of The University of Texas at Arlington in Partial Fulfillment of the Requirements for the Degree of DOCTOR OF PHILOSOPHY THE UNIVERSITY OF TEXAS AT ARLINGTON August 2008 Copyright © by Abraham Miller 2008 All Rights Reserved ACKNOWLEDGEMENTS I am truly indebted to many people for the time and energy they spent in helping reach this goal. Foremost, would be my wife, whom has given me support throughout the entire process; whether it was taking care of certain responsibilities or accompanying me in the field to keep me sane. She has continually encouraged me, offered up ideas and lent a hand anytime I needed it. I cannot put into words how appreciative I am of her. My family has also been supportive in my pursuit and was always interested to learn what I do. In the last months of my dissertation, I have not given them the time they deserve, yet, they have been understanding and supportive of my situation. I would like to thank the graduate students for their input that undoubtedly improved my research and the help that pushed me past the plateaus associated with long-term research. Specifically, I would like to thank Robert Makowsky for his help in the molecular work, Brian Fontenot and John Morse for their help and input in my statistics. I appreciate those who served on my committee, especially my advisor, Dr. Daniel Formanowicz who helped me generate ideas but more importantly helped me focus my ideas and stay on track. -

Scorpiones, Vaejovidae)
Haradon, R. M. 1983. Smeringurus, a new subgenus of Paruroctonus Werner (Scropiones, Vaejovidae). J. Arachnol., 11 :251-270. SMERINGURUS, A NEW SUBGENUS OFPARUROCTONUS WERNER (SCORPIONES, VAEJOVIDAE) Richard M. Haradon 9 High Street Stoneham, Massachusetts 02180 ABSTRACT Paruroctonus Werner consists of two sympatric subgenera, the nominate and Smeringurus, new subgenus. Smeringurus is defined by the presence of numerous short setae on the ventral intercarina l surfaces of metasomal segments I-IV, and by having a significantly more slender metasoma . Smerin- gurus consists of four species endemic to the southwestern deserts of North America : P vachoni Stahnke (type species), P. mesaensis Stahnke, P. grandis (Williams), and P. aridus Soleglad. Two subspecies of P. vachoni are defined by the numbers of primary denticles on the grasping edge of th e pedipalp fingers : P. v. vachoni, from the northern Mojave Desert region, and P. v. immanis Soleglad , new combination, from the central and southern Mojave Desert and the Colorado Desert . Paruroc- tonus vachoni, P, grandis and P. aridus are allopatric, occupying rocky substrates ; each occurs sym - patrically with P. mesaensis, a psammophilous species. INTRODUCTION Within the North American scorpion genus Paruroctonus Werner, 1934, is a distinc t element that includes Paruroctonus vachoni Stahnke, 1961, and four other nominal species. Soleglad (1972) described two of these species, and discussed for the first tim e the distinctive characteristics of the group . Stahnke (1974) doubted the status of both o f Soleglad's species, implicitly, by omitting them from a list of what Stahnke considered t o be the valid Paruroctonus species. The purposes of this report are to evaluate new an d previously employed taxonomic characters, clarify the status of each nominal species, and to propose a subgeneric name for this species group . -

Studies on the Biology and Ecology of the Northern Scorpion, Paruroctonus Boreus (Girard)
Great Basin Naturalist Volume 34 Number 3 Article 1 9-30-1974 Studies on the biology and ecology of the northern scorpion, Paruroctonus boreus (Girard) Gregory Ira Tourtlotte Idaho State University, Pocatello, Idaho Follow this and additional works at: https://scholarsarchive.byu.edu/gbn Recommended Citation Tourtlotte, Gregory Ira (1974) "Studies on the biology and ecology of the northern scorpion, Paruroctonus boreus (Girard)," Great Basin Naturalist: Vol. 34 : No. 3 , Article 1. Available at: https://scholarsarchive.byu.edu/gbn/vol34/iss3/1 This Article is brought to you for free and open access by the Western North American Naturalist Publications at BYU ScholarsArchive. It has been accepted for inclusion in Great Basin Naturalist by an authorized editor of BYU ScholarsArchive. For more information, please contact [email protected], [email protected]. The Great Basin Naturalist Published at Provo, Utah, by Brigham Young University Volume 34 September 30, 1974 No. 3 STUDIES ON THE BIOLOGY AND ECOLOGY OF THE NORTHERN SCORPION, PARUROCTONUS BOREUS (GIRARD)^ Gregory Ira Tourtlotte- Abstract.— An investigation to elucidate some facets of the biology and ecologj' of Paruroctonus boreus (Girard), in southeast Idaho, was conducted from 21 March to 13 October 1972. Three hundred and eighty-five observations were made on 202 scorpions captured in covered pitfalls and by ultraviolet light. The greatest activity occurred at 2130 hours. Surface activity was not observed when ambient temperature was below 10 C, and, as ambient tempera- ture increased, an increase in surface activity was noted. Male scorpions fre- quently moved as much as 30 m between obsei-\'ations, while females seldom moved more than five m. -

Arthropods of Canadian Grasslands
Arthropods of Canadian Grasslands Number 10 2004 Contents Contributions welcome . inside front cover Grasslands project action Ecology and interactions in grassland habitats . 1 Focus sites. 1 Grande grasslands gathering: Dunvegan July 2003 . 2 Grassland project key site 2004: Aweme, Manitoba . 5 Grassland tiger beetles in Canada . 6 The northern scorpion, Paruroctonus boreus, in southern Alberta . 14 Leafhoppers and their relatives from the Canadian Great Plains . 17 Web watch: Build a prairie in the comfort of your own home . 25 Butterfl y inventory of the South Peace River, BC . 26 Update your research projects . 27 Mailing list for the Grasslands Newsletter . 28 Arthropods of Canadian Grasslands supports the grasslands project of the Biological Survey of Canada (Terrestrial Arthropods) by providing information relevant to the study of grassland arthropods in Canada. The Ghost Tiger Beetle, Cicindela lepida Dejean (shown here) may be threatened with local extinction if dune areas shrink. Read about this and other grassland tiger beetles on page 6. 1 Contributions welcome Please consider submitting items to Arthropods of Canadian Grasslands Grassland site Current research – descriptions project reports Short news items Feature articles Grassland species Selected accounts publications Contributions such as these, as well as other items of interest to students of grasslands and their arthropods, are welcomed by the editor. This publication (formerly Newsletter, Arthropods of Canadian Grasslands) appears annually in March; final copy deadline for the next issue is January 31, 2005. Editor: H.V. Danks Biological Survey of Canada (Terrestrial Arthropods) Canadian Museum of Nature P.O. Box 3443, Station “D” Ottawa, ON K1P 6P4 613-566-4787 (tel.) 613-364-4022 (fax) [email protected] Articles without other accreditation are prepared by the Editor. -

Feeding and Excretion in the Scorpion Paruroctonus Mesaensis: Water and Material Balance
J. exp. Biol. 110, 253-265 (1984) 253 gutted in Great Britain © The Company of Biologists Limited 1984 FEEDING AND EXCRETION IN THE SCORPION PARUROCTONUS MESAENSIS: WATER AND MATERIAL BALANCE BY STANLEY D.YOKOTA Department of Biology, University of California at Riverside, Riverside, California 92521, U.SA. Accepted 2 December 1983 SUMMARY Scorpions feed by a process involving the external grinding and digestion of prey, with the ingestion of only the soluble fraction. The water obtained from the prey represents the most important source of water intake for scorpions inhabiting arid regions, placing great importance on the animals' ability to utilize prey water effectively. The scorpionParuroctonus mesaen- sis (Stahnke) was found to ingest a mean of 88 % of the body water of selected prey. However, the scorpion loses 0-37 ml of its own body water per ml of prey water extracted, resulting in a net water gain of 0-51 ml water for every ml of prey water. Fluid uptake by the scorpion has been ascribed to a pharyngeal pumping mechanism. Direct measurements of the suction generated by the pharynx yielded a minimal estimate of its pumping capabil- ity of 130mmHg. The uptake and excretion of nitrogen and electrolytes by Paruroctonus mesaensis on a diet of Tenebrio molitor adults were also analysed. Almost all the potassium ingested was excreted, whereas most of the sodium and chloride were retained, possibly serving to expand haemolymph volume. Assuming a steady state for nitrogen, it was estimated that the net utilizable water obtained from prey, that is the water intake minus the excretory water necessitated by nitrogen excretion, was equivalent to about 35 % of the initial prey water or 69 % of the water ingested. -
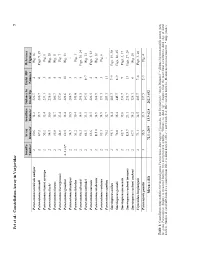
Fet Et Al.: C Onstellation Array in Vaejovidae 7
Fet et al.: Constellation Array in Vaejovidae 7 Sensilla Array Sensillum Distance to Distal MD Reference Number Diameter Diameter Distal Tip Number Figures Paruroctonus arenicola nudipes 2 80.6 14.2 226.7 4 Fig. 16 Paruroctonus arnaudi 2 67.2 15.7 286.7 6 Figs. 9, 29 Paruroctonus bantai saratoga 2 55.2 13.4 231.1 3 Fig. 8 Paruroctonus becki 2 54.5 10.8 238.8 8 Fig. 15 Paruroctonus boreus 2 85.8 15.7 386.7 5 Fig. 11 Paruroctonus borregoensis 2 44.8 13.4 177.8 2 Fig. 7 Paruroctonus gracilior 2–3 (2)* 62.5 15.4 228.6 10 Fig. 13 Paruroctonus hirsutipes 2 34.3 10.5 208.9 3 – Paruroctonus luteolus 2 51.2 10.4 145.0 3 Fig. 1 Paruroctonus silvestrii 2 96.3 14.9 295.5 5 Figs. 10, 24 Paruroctonus stahnkei 2 64.5 13.3 197.5 6–7 Fig. 14 Paruroctonus surensis 2 63.6 12.5 222.5 2 Figs. 5, 23 Paruroctonus utahensis 2 115.0 14.9 288.9 4 Fig. 12 Paruroctonus ventosus 2 85.1 14.9 191.1 3 Fig. 6 Paruroctonus xanthus 2 70.2 12.7 213.3 11 – Smeringurus aridus 2 95.1 14.4 405.7 7–9 Figs. 19, 30 Smeringurus grandis 2 78.8 15.4 445.7 8–9 Figs. 18, 25 Smeringurus mesaensis 2 62.7 12.0 233.7 9 Figs. 2, 17 Smeringurus vachoni immanis 2 128.9 23.1 233.7 13 Figs.