A Comprehensive Molecular Phylogeny of Afrotropical White-Eyes
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Bird List Column A: We Should Encounter (At Least a 90% Chance) Column B: May Encounter (About a 50%-90% Chance) Column C: Possible, but Unlikely (20% – 50% Chance)
THE PHILIPPINES Prospective Bird List Column A: we should encounter (at least a 90% chance) Column B: may encounter (about a 50%-90% chance) Column C: possible, but unlikely (20% – 50% chance) A B C Philippine Megapode (Tabon Scrubfowl) X Megapodius cumingii King Quail X Coturnix chinensis Red Junglefowl X Gallus gallus Palawan Peacock-Pheasant X Polyplectron emphanum Wandering Whistling Duck X Dendrocygna arcuata Eastern Spot-billed Duck X Anas zonorhyncha Philippine Duck X Anas luzonica Garganey X Anas querquedula Little Egret X Egretta garzetta Chinese Egret X Egretta eulophotes Eastern Reef Egret X Egretta sacra Grey Heron X Ardea cinerea Great-billed Heron X Ardea sumatrana Purple Heron X Ardea purpurea Great Egret X Ardea alba Intermediate Egret X Ardea intermedia Cattle Egret X Ardea ibis Javan Pond-Heron X Ardeola speciosa Striated Heron X Butorides striatus Yellow Bittern X Ixobrychus sinensis Von Schrenck's Bittern X Ixobrychus eurhythmus Cinnamon Bittern X Ixobrychus cinnamomeus Black Bittern X Ixobrychus flavicollis Black-crowned Night-Heron X Nycticorax nycticorax Western Osprey X Pandion haliaetus Oriental Honey-Buzzard X Pernis ptilorhynchus Barred Honey-Buzzard X Pernis celebensis Black-winged Kite X Elanus caeruleus Brahminy Kite X Haliastur indus White-bellied Sea-Eagle X Haliaeetus leucogaster Grey-headed Fish-Eagle X Ichthyophaga ichthyaetus ________________________________________________________________________________________________________ WINGS ● 1643 N. Alvernon Way Ste. 109 ● Tucson ● AZ ● 85712 ● www.wingsbirds.com -

Ecological Assessments in the B+WISER Sites
Ecological Assessments in the B+WISER Sites (Northern Sierra Madre Natural Park, Upper Marikina-Kaliwa Forest Reserve, Bago River Watershed and Forest Reserve, Naujan Lake National Park and Subwatersheds, Mt. Kitanglad Range Natural Park and Mt. Apo Natural Park) Philippines Biodiversity & Watersheds Improved for Stronger Economy & Ecosystem Resilience (B+WISER) 23 March 2015 This publication was produced for review by the United States Agency for International Development. It was prepared by Chemonics International Inc. The Biodiversity and Watersheds Improved for Stronger Economy and Ecosystem Resilience Program is funded by the USAID, Contract No. AID-492-C-13-00002 and implemented by Chemonics International in association with: Fauna and Flora International (FFI) Haribon Foundation World Agroforestry Center (ICRAF) The author’s views expressed in this publication do not necessarily reflect the views of the United States Agency for International Development or the United States Government. Ecological Assessments in the B+WISER Sites Philippines Biodiversity and Watersheds Improved for Stronger Economy and Ecosystem Resilience (B+WISER) Program Implemented with: Department of Environment and Natural Resources Other National Government Agencies Local Government Units and Agencies Supported by: United States Agency for International Development Contract No.: AID-492-C-13-00002 Managed by: Chemonics International Inc. in partnership with Fauna and Flora International (FFI) Haribon Foundation World Agroforestry Center (ICRAF) 23 March -

A Philippine Case Study. Environmental Management 2013, 51(6), 1236–1246
Mallari NAD, Collar NJ, McGowan PJK, Marsden SM. Science-driven management of protected areas: a Philippine case study. Environmental Management 2013, 51(6), 1236–1246. Copyright: The final publication is available at Springer via http://dx.doi.org/10.1007/s00267-013-0053-5 DOI link to article: http://dx.doi.org/10.1007/s00267-013-0053-5 Date deposited: 08/04/2016 This work is licensed under a Creative Commons Attribution-NonCommercial 3.0 Unported License Newcastle University ePrints - eprint.ncl.ac.uk 1 Philippine protected areas are not meeting the biodiversity coverage and 2 management effectiveness requirements of Aichi Target 11 3 4 5 Short title: Protected areas and Aichi Target 11 6 7 8 Abstract: Aichi Target 11 of the Convention on Biological Diversity urges, inter alia, that nations 9 protect at least 17% of their land, and that protection is effective and targets areas of importance 10 for biodiversity. Five years before reporting on Aichi targets is due, we assessed the Philippines’ 11 current protected area system for biodiversity coverage, appropriateness of management regimes 12 and capacity to deliver protection. Although protected estate already covers 11% of the 13 Philippines’ land area, 64% of its Key Biodiversity Areas (KBAs) remain unprotected. Few 14 protected areas have appropriate management and governance infrastructures, funding streams, 15 management plans and capacity, and a serious mismatch exists between protected area land 16 zonation regimes and conservation needs of key species. For the Philippines to meet the 17 biodiversity coverage and management effectiveness elements of Aichi Target 11, protected area 18 and KBA boundaries should be aligned, management systems reformed to pursue biodiversity-led 19 targets, and effective management capacity created. -
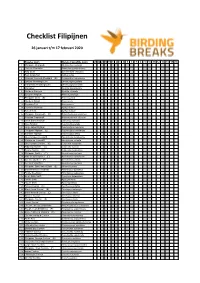
Checklist Filipijnen
Checklist Filipijnen 26 januari t/m 17 februari 2020 Engelse naam Wetenschappelijke naam 27 28 29 30 31 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 1 Philippine Megapode Megapodius cumingii 2 Chinese Francolin Francolinus pintadeanus 3 King Quail Excalfactoria chinensis 4 Red Junglefowl Gallus gallus 5 Palawan Peacock-Pheasant (E) Polyplectron napoleonis 6 Spotted Whistling Duck Dendrocygna guttata 7 Wandering Whistling Duck Dendrocygna arcuata 8 Garganey Spatula querquedula 9 Northern Shoveler Spatula clypeata 10 Eurasian Wigeon Mareca penelope 11 Philippine Duck (E) Anas luzonica 12 Northern Pintail Anas acuta 13 Eurasian Teal Anas crecca 14 Common Pochard Aythya ferina 15 Tufted Duck Aythya fuligula 16 Philippine Frogmouth (E) Batrachostomus septimus 17 Palawan Frogmouth Batrachostomus chaseni 18 Great Eared Nightjar Lyncornis macrotis 19 Grey Nightjar Caprimulgus jotaka 20 Large-tailed Nightjar Caprimulgus macrurus 21 Philippine Nightjar (E) Caprimulgus manillensis 22 Savanna Nightjar Caprimulgus affinis 23 Grey-rumped Treeswift Hemiprocne longipennis 24 Whiskered Treeswift Hemiprocne comata 25 Grey-rumped Swiftlet (E) Collocalia marginata 26 Ridgetop Swiftlet (E) Collocalia isonota 27 Pygmy Swiftlet (E) Collocalia troglodytes 28 Philippine Swiftlet (E) Aerodramus mearnsi 29 Whitehead's Swiftlet (E) Aerodramus whiteheadi 30 Ameline Swiftlet (E) Aerodramus amelis 31 Germain's Swiftlet Aerodramus germani 32 Philippine Spine-tailed Swift (E) Mearnsia picina 33 Brown-backed Needletail Hirundapus giganteus 34 Purple Needletail Hirundapus celebensis -

Ultimate Philippines
The bizarre-looking Philippine Frogmouth. Check those eyes! (Dani Lopez-Velasco). ULTIMATE PHILIPPINES 14 JANUARY – 4/10/17 FEBRUARY 2017 LEADER: DANI LOPEZ-VELASCO This year´s Birdquest “Ultimate Philippines” tour comprised of the main tour and two post-tour extensions, resulting in a five-week endemics bonanza. The first three weeks focused on the better-known islands of Luzon, Palawan and Mindanao, and here we had cracking views of some of those mind-blowing, world´s must-see birds, including Philippine Eagle, Palawan Peacock-Pheasant, Wattled Broadbill and Azure- breasted Pitta, amongst many other endemics. The first extension took us to the central Visayas where exciting endemics such as the stunning Yellow-faced Flameback, the endangered Negros Striped Babbler or the recently described Cebu Hawk-Owl were seen well, and we finished with a trip to Mindoro and remote Northern Luzon, where Scarlet-collared Flowerpecker and Whiskered Pitta delighted us. 1 BirdQuest Tour Report: Ultimate Philippines www.birdquest-tours.com Our success rate with the endemics– the ones you come to the Philippines for- was overall very good, and highlights included no less than 14 species of owl recorded, including superb views of Luzon Scops Owl, 12 species of beautiful kingfishers, including Hombron´s (Blue-capped Wood) and Spotted Wood, 5 endemic racket-tails and 9 species of woodpeckers, including all 5 flamebacks. The once almost impossible Philippine Eagle-Owl showed brilliantly near Manila, odd looking Philippine and Palawan Frogmouths gave the best possible views, impressive Rufous and Writhed Hornbills (amongst 8 species of endemic hornbills) delighted us, and both Scale-feathered and Rough-crested (Red-c) Malkohas proved easy to see. -

A Review of the Recent Advances in the Systematics of the Avian Superfamily Sylvioidea
Chinese Birds 2013, 4(2):99–131 REVIEW DOI 10.5122/cbirds.2013.0016 A review of the recent advances in the systematics of the avian superfamily Sylvioidea Per ALSTRÖM 1,2,, Urban OLSSON 3, Fumin LEI 1 1 Key Laboratory of Zoological Systematics and Evolution, Institute of Zoology, Chinese Academy of Sciences, Beijing 100101, China 2 Swedish Species Information Centre, Swedish University of Agricultural Sciences, Box 7007, SE-750 07 Uppsala, Sweden 3 Systematics and Biodiversity, Department of Biology and Environmental Sciences, University of Gothenburg, Box 463, SE-405 30 Göteborg, Sweden Abstract The systematics of the avian superfamily Sylvioidea are reviewed, focusing on studies of relationships among families and within genera, more superficially on taxonomic studies at the species level. For the families Bernieridae and Phylloscopidae, new analyses based on already published sequence data are presented. Our understanding of relationships has been vastly improved in recent years due to a large number of molecular studies. However, the relationships among the different families remain largely obscured, probably mainly as a result of rapid divergence of the different primary lineages (families). Also, species level taxonomy has been much improved in recent years due to a large number of studies applying molecular markers and/or vocalizations and other life-history data. It seems likely that the number of species will continue to increase, as new groups are being studied with modern integrative methods. Keywords phylogenetic relationships, superfamily Sylvioidea, taxonomy Introduction tanagers, wood-warblers, and icterids). Subsequent studies of DNA sequence data have indicated that both The Passerida was identified by Sibley and Ahlquist Muscicapoidea and Passeroidea, after minor taxonomic (1990) based on DNA-DNA hybridization studies as adjustments, can be recovered as monophyletic (Barker the largest radiation within oscine passerine birds (cf. -

Comprehensive Phylogeny of the Laughingthrushes and Allies (Aves, Leiothrichidae) and a Proposal for a Revised Taxonomy
Received: 13 February 2018 | Revised: 10 April 2018 | Accepted: 1 May 2018 DOI: 10.1111/zsc.12296 ORIGINAL ARTICLE Comprehensive phylogeny of the laughingthrushes and allies (Aves, Leiothrichidae) and a proposal for a revised taxonomy Alice Cibois1 | Magnus Gelang2 | Per Alström3,4,5 | Eric Pasquet6 | Jon Fjeldså7 | Per G. P. Ericson8 | Urban Olsson9 1Natural History Museum of Geneva, Geneva, Switzerland 2Göteborgs Naturhistoriska Museum, Göteborg, Sweden 3Department of Ecology and Genetics, Animal Ecology, Evolutionary Biology Centre, Uppsala University, Uppsala, Sweden 4Swedish Species Information Centre, Swedish University of Agricultural Sciences, Uppsala, Sweden 5Key Laboratory of Zoological Systematics and Evolution, Institute of Zoology, Chinese Academy of Sciences, Beijing, China 6UMS MNHN/CNRS 2700 Outils et Méthodes de la Systématique Intégrative (OMSI) and UMR7205 Institut de Systématique, Evolution, Biodiversité CNRS MNHN UPMC EPHE, Sorbonne Universités, Muséum National d’Histoire Naturelle, Paris, France 7Center for Macroecology, Evolution and Climate, Natural History Museum of Denmark, Zoological Museum, Copenhagen, Denmark 8Department of Vertebrate Zoology, Molecular Systematics Laboratory, Swedish Museum of Natural History, Stockholm, Sweden 9Department of Zoology, University of Gothenburg, Göteborg, Sweden Correspondence Alice Cibois, Natural History Museum Abstract of Geneva, CP 6434, CH 1211 Geneva 6, DNA phylogenies have gradually shed light on the phylogenetic relationships of the Switzerland. large babbler group. We focus in this study on the family Leiothrichidae (laughingth- Email: [email protected] rushes and “song babblers”), which represents the largest clade of babblers in terms Funding information of species diversity. Our phylogeny includes all genera and 82% of the recognized Swedish Research Council, Grant/Award Number: 621-2007-5280 and 2015- species, using mitochondrial and nuclear loci. -

Molecular Systematics of the Avian Superfamily Sylvioidea with Special Regard to the Families Acrocephalidae and Locustellidae
Molecular systematics of the avian superfamily Sylvioidea with special regard to the families Acrocephalidae and Locustellidae (Aves: Passeriformes) I n a u g u r a l d i s s e r t a t i o n zur Erlangung des akademischen Grades doctor rerum naturalium (Dr. rer. nat.) an der Mathematisch-Naturwissenschaftlichen Fakultät der Ernst-Moritz-Arndt-Universität Greifswald vorgelegt von Silke Fregin geboren am 22.05.1976 in Werneck Greifswald, Juni 2012 2 Dekan: Prof. Dr. Klaus Fesser 1. Gutachter : Prof. Dr. Gabriele Uhl 2. Gutachter: Prof. Dr. Staffan Bensch Tag der Promotion: 14.11.2012 3 4 Table of contents Table of contents Zusammenfassung..........................................................................................................................................7 Summary...........................................................................................................................................................9 Abbreviations................................................................................................................................................11 1 General Introduction.....................................................................................................................................13 1.1 DNA-DNA hybridisation studies.........................................................................................................13 1.2 Advantages and disadvantages of DNA sequences..........................................................................14 1.3 Analysis of DNA sequences..................................................................................................................15 -
Remote Philippines
This stunning Whiskered Pitta was, not surprisingly, one of the best birds of the tour. (All photos by DLV, Irene Dy and Brad Hacker). REMOTE PHILIPPINES 3 – 20 / 28 FEBRUARY 2020 LEADER: DANI LOPEZ-VELASCO and IRENE DY With more and more birds being split and ‘available’ in the Philippines, we have resurrected our Remote Philippines tour to give us a great shot at finding a good number of endemics not available on our Classic Philippines tour. The two dovetail perfectly and together really do form the Ultimate Philippines tour. Focusing mostly on the Visayas, Mindoro, the more remote areas of Luzon, followed by a Panay and Tablas extension, the tour was never about a big species total, but purely about seeing those important birds. In total we recorded around 220 species (including half a dozen distinct allospecies), a high percentage of which were ‘diamond’ birds. These included some truly spectacular species, including the gorgeous Whiskered Pitta, which bounded around us at close range, and the critically endangered and very rarely seen Negros Bleeding-heart! Other stunners included the delightful little Negros Scops Owl, the cracking and difficult to find Visayan Broadbill, which we saw just when we were about to leave Bohol, lovely Camiguin, Mindoro, Cebu and Romblon Hawk-Owls, exquisite Dimorphic Dwarf and Northern Silvery Kingfishers, the incomparable Flame-breasted Fruit Dove, the very rare Isabela Oriole, the endangered Walden´s Hornbill and the skulking Bicol Ground Warbler that ran around our feet on our last attempt, to name just a few! 1 BirdQuest Tour Report: Remote Philippines 2020 www.birdquest-tours.com The incomparable Flame-breasted Fruit Dove that showed so well along Infanta Road. -

GRUNDSTEN Philippines 0102
Birding Luzon, Palawan & Mindoro (M. Grundsten, Sweden) 2019 The Philippines, January 27th- February 12th 2019 Luzon – Palawan - Mindoro Front cover Delightful endemic Indigo-banded Kingfisher, Ceyx cyanopectus, at Makiling Botanical Gardens, Los Baños. MG. Participants Måns Grundsten [email protected] (compiler and photos) YouTube: https://www.youtube.com/channel/UChJNc00LXiFfNNnEmvuu2Nw Anders Haglund (photos), Sebbe Nilsson, Claes and Mårten Wikström from Sweden. Tour leader: Bram Demeulemeester, http://www.birdguidingphilippines.com/ Highlights • Fine views of a feeding Flame-breasted Fruit Dove at Mt Polis. • Pre-dawn Bukidnon Woodcock at Mt Polis (previously missed the species at Mount Kitanglad!). • Owling at Subic Bay – including spotlight views of a Slaty-legged Crake. • Beautiful tropical scenery at Sabang, Palawan: Cockatoos in nice evening light. • Legendary male Palawan Peacock-Pheasant at St Paul's. • Many endemics including Falcated Wren-Babbler and Palawan Tit at Sabang. • Smash-and-grab Grey Imperial Pigeon on Pandan Island. • Trek to Mt Victoria through pristine montane forest – Palawan Striped Babbler! • A bag of Mindoro endemics around Lake Libuao, Sablayan Prison and Penal Farm. • Deep voiced and endangered Black-hooded Coucal. • Evening along creek at Sablayan with large flocks of feeding pigeons: Spotted Imperial Pigeon, Yellow- breasted Fruit Dove, Thick-billed Green Pigeon. • Indigo-banded Kingfisher in Makiling Botanical Gardens. • Rare Philippine Eagle Owl at Mt Makiling. • Very last morning with a responsive Whiskered Pitta at Infanta Rd. General information The Philippine Archipelago is among the most endemic-rich areas in the world. And it was sure good to be going back there. At a first glance the itinerary of this trip might seem a bit odd or unusual. -

The Philippines Got Off to a Great Start
Philippines Endangered Endemics 1st May to 18th May 2016 (18 days) Visayan Islands Extension th th 18 to 24 May 2016 (7 days) Palawan Peacock-Pheasant by Sue Wright Trip Report - Philippines Endangered Endemics & Visayan Islands Extension 2016 2 Trip Report compiled by: Forrest Rowland Top Ten Birds of the Tour (as voted by participants): 1. Philippine Eagle 2. Palawan Peacock-Pheasant 3. Azure-breasted Pitta 4. Rufous Hornbill 5. Falcated Wren-Babbler 6. Visayan Broadbill 7. Wattled Broadbill 8. Mantanani Scops-Owl 9. Cebu Hawk-Owl 10. Philippine Frogmouth Our journey through the Philippines got off to a great start. Fortunately, our flight from Manila to Puerto Princesa was a short one and we departed early in the morning, allowing us to hit the ground birding, with plenty of the day left to chalk up an impressive total! We started at a patch of mangroves near the airport where our first birds included our first Philippine Pied Fantails and the aurora subspecies of Olive-backed Sunbird. In the scrub we saw: our first Ashy- fronted Bulbuls, Collared Kingfisher, Rufous-tailed Tailorbird, and Pied Trillers. Our most exciting species near the port was a vibrant Blue-eared Kingfisher perched at eye-level off the boardwalk of our delicious lunch spot! Unfortunately, no Chinese Egrets were in evidence, as it seemed most had left early for breeding grounds - perhaps due to the drought and heat of this El Nino year. This species was one of the astoundingly few dips we had on this productive tour. We began our drive to Sabang, birding a few stops along the way. -

Kennedy Supplement 2016
KENNEDY SUPPLEMENT 2016 WBCP CHECKLIST OF BIRDS OF THE PHILIPPINES 2016 PRESENTED IN THE SEQUENCE AND PAGINATION OF THE KENNEDY GUIDE Highlighting taxonomic and name changes, new records, and newly described species Edited by Christian Perez WBCP Records Committee February 2016 NOTES: 1. All the species listed in the WBCP Checklist of Birds of the Philippines 2016 are shown in the sequence and pagination of “A Guide to the Birds of the Philippines”, by Kennedy et al, 2000, grouped by plate number 2. Species that have not changed are shown in black without any comment. 3. Minor differences such as spelling and removal of the dash are not considered name changes and are not highlighted 4. Taxonomic changes and name changes are highlighted in dark red 5. Species that were split into two or more species are listed in the place of the original species 6. New country records (and one newly described species) are highlighted in blue and shown at the bottom of the relevant plate 7. For resident split species, the range of the new species is indicated (usually the main islands only) to allow identification in the field based on location 8. For migrant split species, a reference is given when possible to a guide for identification in the field PLATE 1 English name in Kennedy Little Grebe Tachybaptus ruficollis Black-necked Grebe Podiceps nigricollis Tahiti Petrel Pseudobulweria rostrata Hawaiian Petrel Pterodroma Dark-rumped Petrel sandwichensis Kermadec Petrel Pterodroma neglecta Bonin Petrel Pterodroma hypoleuca Bulwer's Petrel Bulweria bulwerii