Diving Adaptations of the Cetacean Skeleton Bruno Cozzi*,1, Sandro Mazzariol2, Michela Podestà3 and Alessandro Zotti4
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Human Anatomy (Biology 2) Lecture Notes Updated July 2017 Instructor
Human Anatomy (Biology 2) Lecture Notes Updated July 2017 Instructor: Rebecca Bailey 1 Chapter 1 The Human Body: An Orientation • Terms - Anatomy: the study of body structure and relationships among structures - Physiology: the study of body function • Levels of Organization - Chemical level 1. atoms and molecules - Cells 1. the basic unit of all living things - Tissues 1. cells join together to perform a particular function - Organs 1. tissues join together to perform a particular function - Organ system 1. organs join together to perform a particular function - Organismal 1. the whole body • Organ Systems • Anatomical Position • Regional Names - Axial region 1. head 2. neck 3. trunk a. thorax b. abdomen c. pelvis d. perineum - Appendicular region 1. limbs • Directional Terms - Superior (above) vs. Inferior (below) - Anterior (toward the front) vs. Posterior (toward the back)(Dorsal vs. Ventral) - Medial (toward the midline) vs. Lateral (away from the midline) - Intermediate (between a more medial and a more lateral structure) - Proximal (closer to the point of origin) vs. Distal (farther from the point of origin) - Superficial (toward the surface) vs. Deep (away from the surface) • Planes and Sections divide the body or organ - Frontal or coronal 1. divides into anterior/posterior 2 - Sagittal 1. divides into right and left halves 2. includes midsagittal and parasagittal - Transverse or cross-sectional 1. divides into superior/inferior • Body Cavities - Dorsal 1. cranial cavity 2. vertebral cavity - Ventral 1. lined with serous membrane 2. viscera (organs) covered by serous membrane 3. thoracic cavity a. two pleural cavities contain the lungs b. pericardial cavity contains heart c. the cavities are defined by serous membrane d. -

Module 2 : Anatomy – the Skeleton
Module 2 : Anatomy – The Skeleton In this module you will learn: The functions of the skeletal system The types of bones in the human body The effects of exercise on your bones What happens to the bones as we get older When studying to become a fitness instructor or personal trainer, you will learn all about the anatomy of the human body. Studying the skeleton is one of the foundations of your trade, you will need to know how the body is structured, the names of each bone, types of bones, importance of bone and joint health, detail of the spine and different terms of movement. Without stating the obvious, each of your clients has their own skeleton and you must be fully aware of how this works. This is for many reasons; you are a teacher and must be fully aware of how to prevent injuries, avoid unnecessary stress on the bones and, if qualified, help the client prevent or heal bone and joint related conditions or medical problems. 2.1 Understanding the Skeletal System The skeleton is comprised of 206 different bones that provide 5 main functions: Support mechanism for muscle and tissue Protection for organs Movement with bones, muscles, and joints Storing minerals and blood cells Growth Skeletal System 2.2 Bones are Formed by Ossification Some bones (such as the flat bones of your skull) in the body are formed in a similar stage to connective tissue. The process is known as direct or intramembranous ossification. Other bones are made up of cartilaginous matter, this is developed from future bone in the embryo which then dissolves and is replaced with other bone cells. -

The Muscular System
THE MUSCULAR SYSTEM COMPILED BY HOWIE BAUM 1 Muscles make up the bulk of the body and account for 1/3 of its weight.!! Blood vessels and nerves run to every muscle, helping control and regulate each muscle’s function. The muscular system creates body heat and also moves the: Bones of the Skeletal system Food through Digestive system Blood through the Circulatory system Fluids through the Excretory system MUSCLE TISSUE The body has 3 main types of muscle tissue 1) Skeletal, 2) Smooth, and 3) Cardiac SKELETAL MUSCLE SMOOTH MUSCLE CARDIAC MUSCLE Skeletal muscles attach to and move bones by contracting and relaxing in response to voluntary messages from the nervous system. Skeletal muscle tissue is composed of long cells called muscle fibers that have a striated appearance. Muscle fibers are organized into bundles supplied by blood vessels and innervated by motor neurons. Muscle structure Skeletal (striated or voluntary) muscle consists of densely packed groups of hugely elongated cells known as myofibers. These are grouped into bundles (fascicles). A typical myofiber is 2–3 centimeters ( 3/4–1 1/5 in) long and 0.05millimeters (1/500 inch) in diameter and is composed of narrower structures – myofibrils. These contain thick and thin myofilaments made up mainly of the proteins actin and myosin. Numerous capillaries keep the muscle supplied with the oxygen and glucose needed to fuel contraction. Skeletal Muscles • Skeletal muscles attach to bones by tendons (connective tissue) and enable movement. • Skeletal muscles are mostly voluntary Feel the back of your ankle to feel your Achilles tendon - the largest tendon in your body. -

Is the Skeleton Male Or Female? the Pelvis Tells the Story
Activity: Is the Skeleton Male or Female? The pelvis tells the story. Distinct features adapted for childbearing distinguish adult females from males. Other bones and the skull also have features that can indicate sex, though less reliably. In young children, these sex-related features are less obvious and more difficult to interpret. Subtle sex differences are detectable in younger skeletons, but they become more defined following puberty and sexual maturation. What are the differences? Compare the two illustrations below in Figure 1. Female Pelvic Bones Male Pelvic Bones Broader sciatic notch Narrower sciatic notch Raised auricular surface Flat auricular surface Figure 1. Female and male pelvic bones. (Source: Smithsonian Institution, illustrated by Diana Marques) Figure 2. Pelvic bone of the skeleton in the cellar. (Source: Smithsonian Institution) Skull (Cranium and Mandible) Male Skulls Generally larger than female Larger projections behind the Larger brow ridges, with sloping, ears (mastoid processes) less rounded forehead Square chin with a more vertical Greater definition of muscle (acute) angle of the jaw attachment areas on the back of the head Figure 3. Male skulls. (Source: Smithsonian Institution, illustrated by Diana Marques) Female Skulls Smoother bone surfaces where Smaller projections behind the muscles attach ears (mastoid processes) Less pronounced brow ridges, Chin more pointed, with a larger, with more vertical forehead obtuse angle of the jaw Sharp upper margins of the eye orbits Figure 4. Female skulls. (Source: Smithsonian Institution, illustrated by Diana Marques) What Do You Think? Comparing the skull from the cellar in Figure 5 (below) with the illustrated male and female skulls in Figures 3 and 4, write Male or Female to note the sex depicted by each feature. -

Study Guide Medical Terminology by Thea Liza Batan About the Author
Study Guide Medical Terminology By Thea Liza Batan About the Author Thea Liza Batan earned a Master of Science in Nursing Administration in 2007 from Xavier University in Cincinnati, Ohio. She has worked as a staff nurse, nurse instructor, and level department head. She currently works as a simulation coordinator and a free- lance writer specializing in nursing and healthcare. All terms mentioned in this text that are known to be trademarks or service marks have been appropriately capitalized. Use of a term in this text shouldn’t be regarded as affecting the validity of any trademark or service mark. Copyright © 2017 by Penn Foster, Inc. All rights reserved. No part of the material protected by this copyright may be reproduced or utilized in any form or by any means, electronic or mechanical, including photocopying, recording, or by any information storage and retrieval system, without permission in writing from the copyright owner. Requests for permission to make copies of any part of the work should be mailed to Copyright Permissions, Penn Foster, 925 Oak Street, Scranton, Pennsylvania 18515. Printed in the United States of America CONTENTS INSTRUCTIONS 1 READING ASSIGNMENTS 3 LESSON 1: THE FUNDAMENTALS OF MEDICAL TERMINOLOGY 5 LESSON 2: DIAGNOSIS, INTERVENTION, AND HUMAN BODY TERMS 28 LESSON 3: MUSCULOSKELETAL, CIRCULATORY, AND RESPIRATORY SYSTEM TERMS 44 LESSON 4: DIGESTIVE, URINARY, AND REPRODUCTIVE SYSTEM TERMS 69 LESSON 5: INTEGUMENTARY, NERVOUS, AND ENDOCRINE S YSTEM TERMS 96 SELF-CHECK ANSWERS 134 © PENN FOSTER, INC. 2017 MEDICAL TERMINOLOGY PAGE III Contents INSTRUCTIONS INTRODUCTION Welcome to your course on medical terminology. You’re taking this course because you’re most likely interested in pursuing a health and science career, which entails proficiencyincommunicatingwithhealthcareprofessionalssuchasphysicians,nurses, or dentists. -

Human Biology
KINDERGARTEN HUMAN BIOLOGY 2 WEEKS LESSON PLANS AND ACTIVITIES LIFE CYCLE OVERVIEW OF KINDERGARTEN ORGANISMS WEEK 1. PRE: Comparing large and small organisms. LAB: Classifying and investigating large organisms. POST: Comparing where large animals live. WEEK 2. PRE: Observing how and where organisms live. LAB: Comparing shells. POST: Comparing plants and animals. HUMAN BIOLOGY WEEK 3. PRE: Discovering the different parts of the body. LAB: Discovering and locating parts of the body. POST: Distinguishing the function of external body parts. WEEK 4. PRE: Discovering why bones are important. LAB: Comparing different models of skeletons. POST: Exploring how teeth grow. PLANT LIFE WEEK 5. PRE: Exploring how seeds grow. LAB: Planting seeds. POST: Exploring the uses of plants. WEEK 6. PRE: Defining the parts of a tree and flower. LAB: Comparing seeds and the plants they produce. POST: Comparing flowers, stems, and leaves. NATURAL ENVIRONMENT WEEK 7. PRE: Exploring land and marine environments. LAB: Comparing and contrasting environments. POST: Describing a favorite environment. WEEK 8: PRE: Exploring the order of a natural community. LAB: Dramatizing who eats whom. POST: Characterizing producers and consumers. Math/Science Nucleus ©1990,2000 2 LIFE CYCLE - HUMAN BIOLOGY (KA) PRE LAB Students look at pictures of the body and locate body organs. OBJECTIVES: 1. Discovering organs inside the body. 2. Labeling body parts. VOCABULARY: heart kidney lung muscle organ skeleton stomach MATERIALS: Look Inside Your Body by Gina Ingoglia (Putnam) Human Body Placemats BACKGROUND: A child's body grows every day and unless someone points out the changes, it is difficult for that child to understand what is happening. -

GLOSSARY of MEDICAL and ANATOMICAL TERMS
GLOSSARY of MEDICAL and ANATOMICAL TERMS Abbreviations: • A. Arabic • abb. = abbreviation • c. circa = about • F. French • adj. adjective • G. Greek • Ge. German • cf. compare • L. Latin • dim. = diminutive • OF. Old French • ( ) plural form in brackets A-band abb. of anisotropic band G. anisos = unequal + tropos = turning; meaning having not equal properties in every direction; transverse bands in living skeletal muscle which rotate the plane of polarised light, cf. I-band. Abbé, Ernst. 1840-1905. German physicist; mathematical analysis of optics as a basis for constructing better microscopes; devised oil immersion lens; Abbé condenser. absorption L. absorbere = to suck up. acervulus L. = sand, gritty; brain sand (cf. psammoma body). acetylcholine an ester of choline found in many tissue, synapses & neuromuscular junctions, where it is a neural transmitter. acetylcholinesterase enzyme at motor end-plate responsible for rapid destruction of acetylcholine, a neurotransmitter. acidophilic adj. L. acidus = sour + G. philein = to love; affinity for an acidic dye, such as eosin staining cytoplasmic proteins. acinus (-i) L. = a juicy berry, a grape; applied to small, rounded terminal secretory units of compound exocrine glands that have a small lumen (adj. acinar). acrosome G. akron = extremity + soma = body; head of spermatozoon. actin polymer protein filament found in the intracellular cytoskeleton, particularly in the thin (I-) bands of striated muscle. adenohypophysis G. ade = an acorn + hypophyses = an undergrowth; anterior lobe of hypophysis (cf. pituitary). adenoid G. " + -oeides = in form of; in the form of a gland, glandular; the pharyngeal tonsil. adipocyte L. adeps = fat (of an animal) + G. kytos = a container; cells responsible for storage and metabolism of lipids, found in white fat and brown fat. -
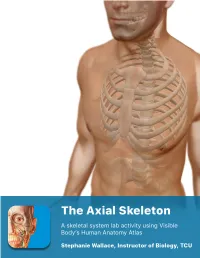
Lab Manual Axial Skeleton Atla
1 PRE-LAB EXERCISES When studying the skeletal system, the bones are often sorted into two broad categories: the axial skeleton and the appendicular skeleton. This lab focuses on the axial skeleton, which consists of the bones that form the axis of the body. The axial skeleton includes bones in the skull, vertebrae, and thoracic cage, as well as the auditory ossicles and hyoid bone. In addition to learning about all the bones of the axial skeleton, it is also important to identify some significant bone markings. Bone markings can have many shapes, including holes, round or sharp projections, and shallow or deep valleys, among others. These markings on the bones serve many purposes, including forming attachments to other bones or muscles and allowing passage of a blood vessel or nerve. It is helpful to understand the meanings of some of the more common bone marking terms. Before we get started, look up the definitions of these common bone marking terms: Canal: Condyle: Facet: Fissure: Foramen: (see Module 10.18 Foramina of Skull) Fossa: Margin: Process: Throughout this exercise, you will notice bold terms. This is meant to focus your attention on these important words. Make sure you pay attention to any bold words and know how to explain their definitions and/or where they are located. Use the following modules to guide your exploration of the axial skeleton. As you explore these bones in Visible Body’s app, also locate the bones and bone markings on any available charts, models, or specimens. You may also find it helpful to palpate bones on yourself or make drawings of the bones with the bone markings labeled. -

Human Anatomy and Physiology
LECTURE NOTES For Nursing Students Human Anatomy and Physiology Nega Assefa Alemaya University Yosief Tsige Jimma University In collaboration with the Ethiopia Public Health Training Initiative, The Carter Center, the Ethiopia Ministry of Health, and the Ethiopia Ministry of Education 2003 Funded under USAID Cooperative Agreement No. 663-A-00-00-0358-00. Produced in collaboration with the Ethiopia Public Health Training Initiative, The Carter Center, the Ethiopia Ministry of Health, and the Ethiopia Ministry of Education. Important Guidelines for Printing and Photocopying Limited permission is granted free of charge to print or photocopy all pages of this publication for educational, not-for-profit use by health care workers, students or faculty. All copies must retain all author credits and copyright notices included in the original document. Under no circumstances is it permissible to sell or distribute on a commercial basis, or to claim authorship of, copies of material reproduced from this publication. ©2003 by Nega Assefa and Yosief Tsige All rights reserved. Except as expressly provided above, no part of this publication may be reproduced or transmitted in any form or by any means, electronic or mechanical, including photocopying, recording, or by any information storage and retrieval system, without written permission of the author or authors. This material is intended for educational use only by practicing health care workers or students and faculty in a health care field. Human Anatomy and Physiology Preface There is a shortage in Ethiopia of teaching / learning material in the area of anatomy and physicalogy for nurses. The Carter Center EPHTI appreciating the problem and promoted the development of this lecture note that could help both the teachers and students. -

Page 1 Human Body Systems: Class Notes Anatomy: Structure of Body Parts
page 1 Human Body Systems: Class Notes Anatomy: Structure of Body Parts. Morphology Physiology: Function of body parts. What they do and how. A & P are very closely related; structure is closely related to function. Characteristics of Life • Movement - self initiated change in position, motion of internal parts • Responsiveness (irritability) - Ability to sense changes within or around the organism and react to them • Growth - increase in body size • Reproduction - Parents produce offspring / producing new individuals • Respiration - Obtaining oxygen (O2), using it to release energy from food substances, and getting rid of wastes • Digestion - Chemically changing (breaking down) food substances, and getting rid of wastes • Absorption - Passage of digested products (food substances) through membranes and into body fluids • Circulation - Movement of substances throughout the body • Assimilation - Changing absorbed substances into chemically different substances • Excretion - Removal of wastes METABOLISM:: All physical and chemical changes occurring in an organism Needs: Water, food, oxygen, heat, pressure. All must be regulated. HOMEOSTASIS: Tendency of the body to maintain a stable, balanced internal environment. Accomplished through self-regulating adjustments. Levels of Organization (from simplest to most complex) Atoms --> Molecules --> Macromolecules --> Organelles --> Cells --> Tissues --> Organs --> Organ Systems --> Organism • A group (mass) of cells working together to carry out certain common functions form a tissue. • A group of tissues working together to carry out certain common functions form an organ. • A group of organs… ORGAN SYSTEMS Body covering. Skin, hair, nails, sweat glands. 1. Integumentary Function: protect underlying tissues and regulate body temperature Bones, ligaments, cartilage 2. Skeletal Function: Support, movement, protection, and production of blood cells Muscles of the body 3. -

!Medical)Terminology) !Anatomy) !Physiology)
!Medical)Terminology) !Anatomy) !Physiology) Jennifer)Figueroa,)M.S.) [email protected]) AAer)this)secBon,)you)will)have) learned…) All)of)the)major)bones)in)the) body…..ROCK)ON)!) )The)major)muscles)in)your)body...) The)parts)of) the)Nervous) System) And)much)much)more)!) Topics) • Medical Terminology • Anatomy and Physiology • Anatomical Terms • Body Systems Medical)Terminology) • Words made from parts – Root: pnea-Breath, arthr-joint – Root with combining form: therm-o + meter = thermometer – Prefix: dys (pain)-pnea, tachy (fast)-pnea – Suffix: arthr-itis (inflammation of), hemophil- iac (pertaining to certain disease) • Compounds of two or more words: smallpox Peri=Around) Cardio=Heart) IBs=)inflammaBon)of) PERICARDITIS?+ Electro=Electric) Myo=Muscle) Gram=recording) ) ELECTROMYOGRAM+(EMG)+ What)about)abbreviaBons?) • ABC’s:)Airway,)Breathing,)CirculaBon) • CVA:)Cerebral)Vascular)Accident) • MVA:)Motor)Vehicle)Accident) • CHF:)CongesBve)Heart)Failure) • CAD:)Coronary)Artery)Disease) ))When)should)you)NOT) ))use)your) Medical)Terminology?) Anatomy)and)Physiology) Anatomical+Posi<on+ Lateral)recumbent)(recovery)) Fowler/Semi!Fowler)PosiBon) PosiBonal)Terms—Trendelenburg) PosiBon) Anatomical)Planes) Transverse)Plane=Horizontal)Plane=X)Sec) Frontal)Plane=)Coronal)Plane) Oblique)plane) DirecBonal)Terms) Lateral+ Medial+ Anterior+ Posterior+ Superior/Rostral+ Palmar) Ventral+ Dorsal+ Distal+ Proximal+ Plantar) Inferior/Caudal+ Anatomical Position A is _____ to B. A is _____ to B. A is _____ to B. A is _____ and ______ to B. A is _____ and -

(Frontal Sinus, Coronal Suture) Parietal Bone
Axial Skeleton Skull (cranium) Frontal Bone (frontal sinus, coronal suture) Parietal Bone (sagittal suture) Sphenoid Bone (sella turcica, sphenoid sinus) Temporal Bone (mastoid process, styloid process, external auditory meatus) malleus incus stapes Occipital Bone (occipital condyles, foramen magnum) Ethmoid Bone (nasal conchae, cribriform plate, crista galli, ethmoid sinus, perpendicular plate) Lacrimal Bone Zygomatic Bone Maxilla Bone (hard palate, palatine process, maxillary sinus) Palatine Bone Nasal Bone Vomer Bone Mandible Hyoid Bone Vertebral Column (general markings: body, vertebral foramen, transverse process, spinous process, superior and inferior articular processes) Cervical Vertebrae (transverse foramina) Atlas (absence of body, "yes" movement) Axis (dens, "no" movement) Thoracic Vertebrae (facets on body and transverse processes) Lumbar Vertebrae (largest) Sacral Vertebrae 5 fused vertebrae) Coccyx (3 to 5 vestigial vertebrae, body only on most) Bony Thorax Ribs (costal cartilage, true ribs, false ribs, floating ribs, facets) Sternum Manubrium Body Xiphoid Process Appendicular Skeleton Upper Limb Pectoral Girdle Scapula (acromion, coracoid process, glenoid cavity, spine) Clavicle Upper Arm Humerus (head, greater tubercle, lesser tubercle, olecranon fossa) Forearm Radius Ulna (olecranon process) Hand Carpals Metacarpals Phalanges Lower Limb Pelvic Girdle Os Coxae (sacroiliac joint, acetabulum, obturator foramen, false pelvis, difference between male and female pelvis) Ilium (iliac crest) Ischium (ischial tuberosity) Pubis (pubic symphysis) Thigh Femur (head, neck) Patella Lower Leg Tibia Fibula Foot Tarsals Metatarsals Phalanges.