Microrna-146 Protects A549 and H1975 Cells from LPS-Induced Apoptosis and Inflammation Injury
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Report 2011–5010
Shahejie−Shahejie/Guantao/Wumishan and Carboniferous/Permian Coal−Paleozoic Total Petroleum Systems in the Bohaiwan Basin, China (based on geologic studies for the 2000 World Energy Assessment Project of the U.S. Geological Survey) 114° 122° Heilongjiang 46° Mongolia Jilin Nei Mongol Liaoning Liao He Hebei North Korea Beijing Korea Bohai Bay Bohaiwan Bay 38° Basin Shanxi Huang He Shandong Yellow Sea Henan Jiangsu 0 200 MI Anhui 0 200 KM Hubei Shanghai Scientific Investigations Report 2011–5010 U.S. Department of the Interior U.S. Geological Survey Shahejie−Shahejie/Guantao/Wumishan and Carboniferous/Permian Coal−Paleozoic Total Petroleum Systems in the Bohaiwan Basin, China (based on geologic studies for the 2000 World Energy Assessment Project of the U.S. Geological Survey) By Robert T. Ryder, Jin Qiang, Peter J. McCabe, Vito F. Nuccio, and Felix Persits Scientific Investigations Report 2011–5010 U.S. Department of the Interior U.S. Geological Survey U.S. Department of the Interior KEN SALAZAR, Secretary U.S. Geological Survey Marcia K. McNutt, Director U.S. Geological Survey, Reston, Virginia: 2012 For more information on the USGS—the Federal source for science about the Earth, its natural and living resources, natural hazards, and the environment, visit http://www.usgs.gov or call 1–888–ASK–USGS. For an overview of USGS information products, including maps, imagery, and publications, visit http://www.usgs.gov/pubprod To order this and other USGS information products, visit http://store.usgs.gov Any use of trade, product, or firm names is for descriptive purposes only and does not imply endorsement by the U.S. -

Original Article Effect of Multidimensional Comprehensive Intervention on Medication Compliance, Social Function and Incidence of MACE in Patients Undergoing PCI
Am J Transl Res 2021;13(7):8058-8066 www.ajtr.org /ISSN:1943-8141/AJTR0131043 Original Article Effect of multidimensional comprehensive intervention on medication compliance, social function and incidence of MACE in patients undergoing PCI Yan Zhang1*, Yuhua Yang2*, Jinggang Xiao3, Yao Sun4, Suping Yang5, Xintao Fu6 1Department of Cardiovascular Medicine, Jinan People’s Hospital Affiliated to Shandong First Medical University, Jinan, Shandong, China; 2Department of Drug Dispensing, Zibo Central Hospital, Zibo, Shandong, China; 3The Second Department of Cardiovascular Medicine, Linqing People’s Hospital, Linqing, Shandong, China; 4Depart- ment of General Practice, Zibo Central Hospital, Zibo, Shandong, China; 5Department of Geriatrics, Binzhou Hos- pital of Traditional Chinese Medicine, Binzhou, Shandong, China; 6Department of Cardiac Surgery, Zibo Municipal Hospital, Zibo, Shandong, China. *Equal contributors and co-first authors. Received January 29, 2021; Accepted March 29, 2021; Epub July 15, 2021; Published July 30, 2021 Abstract: Objective: To analyze the effect of multidimensional comprehensive intervention on medication compli- ance, social function and incidence of major adverse cardiovascular events (MACE) in patients undergoing percuta- neous coronary intervention (PCI). Methods: Ninety-eight patients with coronary heart disease (CHD) who underwent PCI in our hospital were selected and divided into the regular group (n=46, receiving regular nursing intervention) and the comprehensive group (n=52, receiving multidimensional comprehensive nursing intervention) according to the different nursing intervention methods. The medication compliance, social function, quality of life, and inci- dence of MACE were compared between the two groups. Results: The comprehensive group showed significantly higher rates of taking medication on time, taking medication according to the proper amount, taking medication at the recomended times, no increase or decrease in the amount of medication, and taking medication without interruption than the regular group (P < 0.05). -

Information Extraction of Tourist Geological Resources Based on 3D Visualization Remote Sensing Image
The International Archives of the Photogrammetry, Remote Sensing and Spatial Information Sciences, Volume XLII-3, 2018 ISPRS TC III Mid-term Symposium “Developments, Technologies and Applications in Remote Sensing”, 7–10 May, Beijing, China INFORMATION EXTRACTION OF TOURIST GEOLOGICAL RESOURCES BASED ON 3D VISUALIZATION REMOTE SENSING IMAGE Wang Xulong Shandong Remote Sensing Center of Technology and Application, Jinan, China [email protected] Commission III, WG III/1 KEY WORDS: Remote sensing image, 3D visualization, Tourism geological resources, Information Extraction, Investigation ABSTRACT: Tourism geological resources are of high value in admiration, scientific research and universal education, which need to be protected and rationally utilized. In the past, most of the remote sensing investigations of tourism geological resources used two-dimensional remote sensing interpretation method, which made it difficult for some geological heritages to be interpreted and led to the omission of some information. This aim of this paper is to assess the value of a method using the three-dimensional visual remote sensing image to extract information of geological heritages. skyline software system is applied to fuse the 0.36m aerial images and 5m interval DEM to establish the digital earth model. Based on the three-dimensional shape, color tone, shadow, texture and other image features, the distribution of tourism geological resources in Shandong Province and the location of geological heritage sites were obtained, such as geological structure, DaiGu landform, granite landform, Volcanic landform, sandy landform, Waterscapes, etc. The results show that using this method for remote sensing interpretation is highly recognizable, making the interpretation more accurate and comprehensive. 1. INTRODUCTION features of the geological heritage displayed in the three- dimensional visualized remote sensing image, remote sensing Tourism geological resources are of high value in admiration, interpretation keys are established(Zhao, 2013). -

2012 Summarized Annual Report of Qilu Bank Co., Ltd. (The Annual Report Is Prepared in Chinese and English
2012 Summarized Annual Report of Qilu Bank Co., Ltd. (The annual report is prepared in Chinese and English. English translation is purely for reference only. Should there be any inconsistencies between them; the report in Chinese shall prevail.) Ⅰ. General Introduction ()Ⅰ Legal Name in Chinese:齐鲁银行股份有限公司 (Abbreviation: 齐鲁银行 ) Legal Name in English: QiLu Bank Co., Ltd. (Ⅱ ) Legal Representative: Wang Xiaochun (Ⅲ ) Secretary of the Board of Directors: Mao Fangzhu Address: No.176 Shunhe Street, Shizhong District, Jinan City, Shandong Province Tel: 0531-86075850 Fax: 0531-86923511 Email: [email protected] (Ⅳ ) Registered Address: No.176 Shunhe Street, Shizhong District, Jinan City Office Address: No.176 Shunhe Street, Shizhong District, Jinan City Postcode: 250001 Website: http://www.qlbchina.com (Ⅴ ) Newspapers for Information Disclosure: Financial News Website for Information Disclosure: http://www.qlbchina.com Place where copies of the annual report are available at: The Board of Directors' Office of the Bank (Ⅵ ) Other Relevant Information Date of the Initial Registration: 5 June 1996 Address of the Initial Registration: Shandong Administration for Industry and Commerce Corporate Business License Number: 370000018009391 Tax Registration Number: Ludishuiji Zi No.370103264352296 Auditors: Ernst &Young Hua Ming LLP Auditors’ Address: Level 16, Ernst & Young Tower, Oriental Plaza No.1, East Changan Avenue, Dong Cheng District, Beijing, China 1 II. Financial Highlights (I) Main Profit Indicators for the Reporting Period (Group) Unit -

Analysis of Spatial-Temporal Distribution of Notifiable Respiratory
Li et al. BMC Public Health (2021) 21:1597 https://doi.org/10.1186/s12889-021-11627-6 RESEARCH ARTICLE Open Access Analysis of spatial-temporal distribution of notifiable respiratory infectious diseases in Shandong Province, China during 2005– 2014 Xiaomei Li1†, Dongzhen Chen1,2†, Yan Zhang3†, Xiaojia Xue4, Shengyang Zhang5, Meng Chen6, Xuena Liu1* and Guoyong Ding1* Abstract Background: Little comprehensive information on overall epidemic trend of notifiable respiratory infectious diseases is available in Shandong Province, China. This study aimed to determine the spatiotemporal distribution and epidemic characteristics of notifiable respiratory infectious diseases. Methods: Time series was firstly performed to describe the temporal distribution feature of notifiable respiratory infectious diseases during 2005–2014 in Shandong Province. GIS Natural Breaks (Jenks) was applied to divide the average annual incidence of notifiable respiratory infectious diseases into five grades. Spatial empirical Bayesian smoothed risk maps and excess risk maps were further used to investigate spatial patterns of notifiable respiratory infectious diseases. Global and local Moran’s I statistics were used to measure the spatial autocorrelation. Spatial- temporal scanning was used to detect spatiotemporal clusters and identify high-risk locations. Results: A total of 537,506 cases of notifiable respiratory infectious diseases were reported in Shandong Province during 2005–2014. The morbidity of notifiable respiratory infectious diseases had obvious seasonality with high morbidity in winter and spring. Local Moran’s I analysis showed that there were 5, 23, 24, 4, 20, 8, 14, 10 and 7 high-risk counties determined for influenza A (H1N1), measles, tuberculosis, meningococcal meningitis, pertussis, scarlet fever, influenza, mumps and rubella, respectively. -

Cangzhou Beijing Tongzhou Tianjin Dezhou Linqing Liaocheng Ji'ning
Part II Waterway Scaling on Regions Tongzhou Tianjin Cangzhou Dezhou Beijing 165 Linqing Liaocheng Ji’ning Taier zhuang Xuzhou Suqian Huaiyin Zhenjiang Yangzhou Changzhou Hangzhou Suzhou Wuxi Jiaxing 0 5km Figure 7-1 Urban forms of waterway and 18 cities in the early 20th century. Waterway Scaling Regional Development—— A Cultural Landscape Perspective in China Grand Canal Tianjin Cangzhou Beijing Tongzhou Dezhou 166 Linqing Liaocheng Ji’ning Taier zhuang Xuzhou Suqian Huaiyin Yangzhou Zhenjiang Changzhou Suzhou Wuxi Jiaxing Hangzhou Figure 7-2 Prototypes of waterway and 18 cities in the early 20th century. Part II Waterway Scaling on Regions different water values for cities, such as ecological, recreational, transportation, etc. Compare representative 18 cities of canal, we Chapter 7 Regions of Waterway could understand the way how the city was treating its waterway. I analyzed that a pre-industrial walled city could have five typical relationships to the 7.1 Introduction waterway, and now it would leave only one simple pattern. Waterway is a corridor working on the spatial regional and metropolitan plan. A diachronic analysis tries to explain regions of waterway by two periods: the first period is when This chapter described the historical and modern the Grand Canal belonged to the empire, but none Grand Canal in its cities, how the waterway working of the walled cities could open their door to this as one articulation part in ancient city and modern waterway excepted the capital Beijing; the second 167 metropolis areas, even though some of the waterways period is when cities have expanded to a larger urban have lost navigable capacity. -
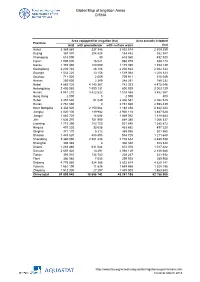
Global Map of Irrigation Areas CHINA
Global Map of Irrigation Areas CHINA Area equipped for irrigation (ha) Area actually irrigated Province total with groundwater with surface water (ha) Anhui 3 369 860 337 346 3 032 514 2 309 259 Beijing 367 870 204 428 163 442 352 387 Chongqing 618 090 30 618 060 432 520 Fujian 1 005 000 16 021 988 979 938 174 Gansu 1 355 480 180 090 1 175 390 1 153 139 Guangdong 2 230 740 28 106 2 202 634 2 042 344 Guangxi 1 532 220 13 156 1 519 064 1 208 323 Guizhou 711 920 2 009 709 911 515 049 Hainan 250 600 2 349 248 251 189 232 Hebei 4 885 720 4 143 367 742 353 4 475 046 Heilongjiang 2 400 060 1 599 131 800 929 2 003 129 Henan 4 941 210 3 422 622 1 518 588 3 862 567 Hong Kong 2 000 0 2 000 800 Hubei 2 457 630 51 049 2 406 581 2 082 525 Hunan 2 761 660 0 2 761 660 2 598 439 Inner Mongolia 3 332 520 2 150 064 1 182 456 2 842 223 Jiangsu 4 020 100 119 982 3 900 118 3 487 628 Jiangxi 1 883 720 14 688 1 869 032 1 818 684 Jilin 1 636 370 751 990 884 380 1 066 337 Liaoning 1 715 390 783 750 931 640 1 385 872 Ningxia 497 220 33 538 463 682 497 220 Qinghai 371 170 5 212 365 958 301 560 Shaanxi 1 443 620 488 895 954 725 1 211 648 Shandong 5 360 090 2 581 448 2 778 642 4 485 538 Shanghai 308 340 0 308 340 308 340 Shanxi 1 283 460 611 084 672 376 1 017 422 Sichuan 2 607 420 13 291 2 594 129 2 140 680 Tianjin 393 010 134 743 258 267 321 932 Tibet 306 980 7 055 299 925 289 908 Xinjiang 4 776 980 924 366 3 852 614 4 629 141 Yunnan 1 561 190 11 635 1 549 555 1 328 186 Zhejiang 1 512 300 27 297 1 485 003 1 463 653 China total 61 899 940 18 658 742 43 241 198 52 -

Official-Gazette-Vol.-89.Pdf
ສາທາລະນະລດັ ປະຊາທິປະໄຕ ປະຊາຊນົ ລາວ LAO PEOPLE’S DEMOCRATIC REPUBLIC ກະຊວງ ວທິ ະຍາສາດ ແລະ ເຕັກໂນໂລຊ ີ ກມົ ຊບັ ສນິ ທາງປນັ ຍາ MINISTRY OF SCIENCE AND TECHNOLOGY DEPARTMENT OF INTELLECTUAL PROPERTY ຈດົ ໝາຍເຫດທາງລດັ ຖະການ ກຽ່ ວກບັ ການເຜີຍແຜຜ່ ນົ ຂອງການຈດົ ທະບຽນ ຊບັ ສນິ ອດຸ ສາຫະກາ ຢ່ ສປປ ລາວ OFFICIAL GAZETTE OF INDUSTRIAL PROPERTY ສະບບັ ທີ (Vol.) 89 Official Gazette of Industrial Property Vol. 89, 20/03/2020 ສາລະບານ ພາກທີ I: ຜນົ ຂອງການຂໍຈດົ ທະບຽນເຄື່ ອ ງໝາຍການຄາ້ ພາກທີ II: ຜນົ ຂອງການຈດົ ທະບຽນເຄື່ ອ ງໝາຍການຄາ້ ພາກທີ III: ຜນົ ຂອງການຂໍຈດົ ທະບຽນເຄື່ ອ ງໝາຍການຄາ້ ສາກນົ ພາກທີ IV: ຜນົ ຂອງການຈດົ ທະບຽນເຄື່ ອ ງໝາຍການຄາ້ ສາກນົ ພາກທີ V: ຜນົ ຂອງການຕື່ ອາຍກຸ ານຈດົ ທະບຽນເຄື່ ອ ງໝາຍການຄາ້ CONTENT Part I: New Application of Trademark Part II: Registration of Trademark Part III: Madrid New Application of Trademark Part IV: Madrid Registration of Trademark Part V: Renewal of Trademark I Official Gazette of Industrial Property Vol. 89, 20/03/2020 ພາກທີ I ຜນົ ຂອງການຂໍຈດົ ທະບຽນເຄື່ ອ ງໝາຍການຄາ້ ຄາ ແນະນາ ກຽື່ ວກບັ ລະຫດັ ຫຍ ້ (540) ເຄື່ ອ ງໝາຍການຄາ້ (511) ໝວດຂອງສນິ ຄາ້ ແລະ ການບ ລິການ (210) ເລກທີຄາ ຮອ້ ງ (220) ວນັ ທີທື່ ີຍື່ ນ ຄາ ຮອ້ ງ (732) ຊື່ ແລະ ທື່ ີຢື່ ຂອງຜຍ້ ື່ ນຄາ ຮອ້ ງ Part I New Application of Trademark Introduction of Codes (540) Trademark (511) Classification of goods and services (Nice Classification) (210) Number of the application (220) Date of filing of the application (732) Name and Address of applicant II Official Gazette of Industrial Property Vol. 89, 20/03/2020 ພາກທີ II ຜນົ ຂອງການຈດົ ທະບຽນເຄື່ ອ ງໝາຍການຄາ້ ຄາ ແນະນາ ກຽື່ ວກບັ ລະຫດັ ຫຍ ້ (540) ເຄື່ -

2010 Summarized Annual Report of Qilu Bank Co., Ltd. (The Annual Report Is Prepared in Chinese and English
2010 Summarized Annual Report of Qilu Bank Co., Ltd. (The annual report is prepared in Chinese and English. English translation is purely for reference only. Should there be any inconsistencies between them; the report in Chinese shall prevail.) Ⅰ. General Introduction ()Ⅰ Legal Name in Chinese:齐鲁银行股份有限公司 (Abbreviation: 齐鲁银行 ) Legal Name in English: Qilu Bank Co., Ltd. (Ⅱ ) Legal Representative: Qiu Yunzhang (Ⅲ ) Secretary of the Board of Directors: Mao Fangzhu Address: No.176 Shunhe Street, Shizhong District, Jinan City, Shandong Province Tel: 0531-86075850 Fax: 0531-86923511 Email: [email protected] (Ⅳ ) Registered Address: No.176 Shunhe Street, Shizhong District, Jinan City Office Address: No.176 Shunhe Street, Shizhong District, Jinan City Postcode: 250001 Website: http://www.qlbchina.com ()Ⅴ Place where copies of the annual report are available at: The Board of Directors' Office of the Bank (Ⅵ ) Other Relevant Information Date of the Initial Registration: 5 June 1996 Address of the Initial Registration: Shandong Administration for Industry and Commerce Corporate Business License Number: 370000018009391 Tax Registration Number: Ludishuiji Zi No.370103264352296 Auditors: Ernst & Young Hua Ming LLP Auditors’ Address: Level 16, Ernst & Young Tower, Oriental Plaza No.1, East Changan Avenue, Dong Cheng District, Beijing, China 1 II. Financial Highlights (I) Main Profit Indicators for the Reporting Period Unit (RMB thousand) Item Amount Profit Before Tax 396,813 Net Profit 333,382 Operating Income 2,301,954 Operating Expense 1,909,866 -

Qingdao Port International Co Annual Report 12748B.Pdf
CONTENTS PAGE DEFINITIONS 2 CORPORATE INFORMATION 6 COMPANY PROFILE 8 2017 MAJOR EVENTS 12 FINANCIAL HIGHLIGHTS 17 CHAIRMAN’S STATEMENT 19 MANAGEMENT DISCUSSION AND ANALYSIS 22 DIRECTORS, SUPERVISORS AND SENIOR MANAGEMENT 44 DIRECTORS’ REPORT 51 SUPERVISORS’ REPORT 68 CORPORATE GOVERNANCE REPORT 71 AUDITOR’S REPORT 85 CONSOLIDATED AND COMPANY BALANCE SHEET 91 CONSOLIDATED AND COMPANY INCOME STATEMENT 95 CONSOLIDATED AND COMPANY CASH FLOW STATEMENT 98 CONSOLIDATED STATEMENT OF CHANGES IN SHAREHOLDERS’ EQUITY 102 COMPANY STATEMENT OF CHANGES IN SHAREHOLDERS’ EQUITY 104 NOTES TO THE FINANCIAL STATEMENTS 106 DEFINITIONS Unless the context otherwise requires, the following expressions shall have the following meanings: “2017 AGM” the annual general meeting for the year 2017 of the Company to be held on 6 June 2018 “Articles of Association” the articles of association of the Company, as amended from time to time “Board” the board of directors of Qingdao Port International Co., Ltd. (青島港國際股份有限公司) “CFS” container freight station, of which, container freight station at loading ports refers to the location designated by carriers for the receiving of cargo to be loaded into containers by the carrier, while container freight station at discharge or destination ports refer to the location designated by carriers for de-vanning of containerized cargo “Company” Qingdao Port International Co., Ltd. (青島港國際股份有限公司), a joint stock company established in the PRC with limited liability on 15 November 2013 “Consolidated Group Companies” the Company (including -

Spatio-Temporal Evolution of Economic Polycentric Pattern at County Level in Shandong Province
E3S Web of Conferences 300, 02017 (2021) https://doi.org/10.1051/e3sconf/202130002017 ICEPESE2021 Spatio-temporal evolution of economic polycentric pattern at county level in Shandong Province Fan Wu, Jun Chang*, and Lifei Li College of Geography and Environment, Shandong Normal University, 250358 Jinan, China Abstract. From the perspective of economy and comprehensive development level, this study used the gravity model, spatial autocorrelation analysis and principal component analysis to quantitatively measure the spatiotemporal evolution pattern of multi-centers at county level in Shandong Province. The results show that the economic ties among counties in Shandong Province are getting closer and closer. By 2016, Jinan-Zibo-Qingdao and Jining, Zaozhuang have basically formed three strong economic ties. The amount of counties with high-high GDP and low-low GDP are decreasing, while the amount of counties with low- high GDP are increasing. The gap between the density of output value and the level of economic development is narrowing, showing a trend of multi- center development. In the future development, Shandong Province should strengthen the integration of resources within the province, form a reasonable industrial division of labor, strengthen the cooperation among enterprises, promote the regional integration construction, and realize the multi-center spatial development model of cooperation. Keywords. Economic polycentric pattern, spatio-temporal evolution, county level, Shandong Province. 1 Introduction With the strengthening of economic globalization and the advancement of urbanization, as the product of regional high industrialization and urbanization, polycentric urban area has gradually replaced the city and become the basic regional unit participating in international competition and division of labor [1]. -

Finding Carbon Financing for Landfill Projects
FindingFinding CarbonCarbon FinanceFinance forfor LandfillLandfill Projects:Projects: OngoingOngoing PilotPilot ActivitiesActivities inin ShandongShandong ProvinceProvince Methane to Markets Partnership Expo Beijing, October 31, 2007 Vinay Deodhar, CDM Specialist & Team Leader CMI-TSF Li Bo, Chief Coordinator SPPMO OutlineOutline • Funding gap and project finance for LF projects • ADB role in carbon finance • Case study of the Shandong LFG project FundingFunding GapGap inin LFLF ProjectsProjects • The main stakeholders for LF projects - City governments, who have the primary responsibility of waste management • The service is generally financed from the collection of property tax • However, many other priorities use up the property tax collection • As a result, Solid Waste Management is left inadequately funded • Preferred financing model is increasingly public- private-partnership (PPP) ProjectProject financefinance forfor LFLF projectsprojects • The PPP model works on off balance sheet terms • Tipping fees being the revenue stream and selection criteria for selection of private partner in the bidding process • In this process carbon finance is a worthy addition to make the projects investment grade • LFG capture and utilization projects satisfy all the eligibility criteria of CDM methodology • The CDM revenues for such projects helps in removing major investment barrier ADBADB rolerole inin carboncarbon financefinance • Provide underlying finance – Loan, equity, guarantees • Through REACH and CDM Facility ADB provided capacity building