Coyote (Canis Latrans) Occurrence Relative to Human Use on Glenbow Ranch Provincial Park, Alberta
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
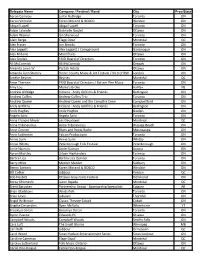
Aaron Comeau Justin Rutledge Toronto ON
Delegate Name Company / Festival / Band City Prov/State Aaron Comeau Justin Rutledge Toronto ON Aaron Verhulst Karen Morand & BOSCO Windsor ON Abigail Lapell Abigail Lapell Toronto ON Adam Lalonde Gabrielle Goulet Ottawa ON Adam Warner Ian Sherwood Toronto ON Alain Berge Élage Diouf Montréal QC Alec Fraser Jon Brooks Toronto ON Alex Leggett Alex Leggett / Campground Gananoque ON Alex Millaire Moonfruits Ottawa ON Alex Sinclair FMO Board of Directors Toronto ON Ali McCormick Ali McCormick Ompah ON Amanda Lowe W. Partick Artists Ottawa ON Amanda Lynn Stubley Home County Music & Art Festival / 94.9 CHRW London ON Amélie Beyries Beyries Montréal QC Amie Therrien FMO Board of Directors / Balsam Pier Music Toronto ON Amy Lou Mama's Broke Halifax NS Andrew Aldridge Kidzent - Andy Griffiths & Friends Burlington ON Andrew Collins Andrew Collins Trio Toronto ON Andrew Queen Andrew Queen and the Campfire Crew Campbellford ON Andy Griffiths Kidzent - Andy Griffiths & Friends Burlington ON Andy Hughes Andy Hughes Guelph ON Angela Saini Angela Saini Toronto ON Anna Frances Meyer Les Deuxluxes Montreal QC Anna Tribinevicius Anna Tribinevicius Wasaga Beach ON Anne Connor Blues and Roots Radio Mississauga ON Anne Lederman Falcon Productions Toronto ON Annie Sumi Annie Sumi Whitby ON Annie Whitty Peterborough Folk Festival Peterborough ON Arnie Naiman Arnie Naiman Aurora ON Ayron Mortley Urban Highlanders Toronto ON Barbra Lica Barbra Lica Quintet Toronto ON Barry Miles Murder Murder Sudbury ON Benny Santoro Karen Morand & BOSCO Windsor ON Bill Collier -

Environmental Commission Agenda Report
ENVIRONMENTAL COMMISSION AGENDA REPORT DATE: AUGUST 25, 2011 TO: HONORABLE CHAIR AND MEMBERS OF THE ENVIRONMENTAL COMMISISON FROM: ALEX FARASSATI, PH.D., ENVIRONMENTAL SERVICES SUPERVISOR SUBJECT: COYOTE MANAGEMENT IN CALABASAS MEETING DATE: SEPTEMBER 6, 2011 SUMMARY RECOMMENDATION: That the Environmental Commission review and discuss this report and recommend a course of action to the City Council for consideration. BACKGROUND: During the public comment section of a City Council meeting held on July 13, 2011, several Calabasas residents and wildlife activists came out in opposition of the cities policy on trapping coyotes. Since this meeting all coyote trappings have been suspended and all homeowners associations have been notified of the suspension, until the Environmental Commission had the opportunity to discuss the issue and make a recommendation to the City Council. Meanwhile an on-line petition was initiated by Project Coyote that was signed by 1,981 individuals throughout the world requesting permanent suspension of coyote trapping within the city. Prior to suspension, the Calabasas coyote management procedure has followed Los Angeles County protocol described below: Environmental Commission Agenda Report September 6, 2011 1. A Calabasas homeowner contacts Public Works Landscape Division staff with a request for coyote abatement; 2. During the conversation with the homeowner, by utilizing GIS software imaging, Public Works Landscape Division staff determines if there is public open space or common area land adjacent to the homeowner’s parcel, where a County trapper might be able to safely place a trap; 3. If there is, Public Works Landscape Division staff contacts the Los Angeles County, Agricultural Commissioner/Weights and Measures, Pest Management Division and gives them permission to have one of their trappers contact the homeowner, visit the site, and determine if a trap can safely be placed; 4. -

1944 Wolf Attacks on Humans: an Update for 2002–2020
1944 Wolf attacks on humans: an update for 2002–2020 John D. C. Linnell, Ekaterina Kovtun & Ive Rouart NINA Publications NINA Report (NINA Rapport) This is NINA’s ordinary form of reporting completed research, monitoring or review work to clients. In addition, the series will include much of the institute’s other reporting, for example from seminars and conferences, results of internal research and review work and literature studies, etc. NINA NINA Special Report (NINA Temahefte) Special reports are produced as required and the series ranges widely: from systematic identification keys to information on important problem areas in society. Usually given a popular scientific form with weight on illustrations. NINA Factsheet (NINA Fakta) Factsheets have as their goal to make NINA’s research results quickly and easily accessible to the general public. Fact sheets give a short presentation of some of our most important research themes. Other publishing. In addition to reporting in NINA's own series, the institute’s employees publish a large proportion of their research results in international scientific journals and in popular academic books and journals. Wolf attacks on humans: an update for 2002– 2020 John D. C. Linnell Ekaterina Kovtun Ive Rouart Norwegian Institute for Nature Research NINA Report 1944 Linnell, J. D. C., Kovtun, E. & Rouart, I. 2021. Wolf attacks on hu- mans: an update for 2002–2020. NINA Report 1944 Norwegian In- stitute for Nature Research. Trondheim, January, 2021 ISSN: 1504-3312 ISBN: 978-82-426-4721-4 COPYRIGHT © Norwegian -

Biolink April, 2010
Volume 46 (1) BioLink April, 2010 The Official Newsletter of the Atlantic Society of Fish and Wildlife Biologists Join us and celebrate Earth Day, April 22, 2010 ,0930-1530h for the ASFWB Spring Seminar titled “Aboriginal Perspectives on Fish & Wildlife Management". It will be hosted at the Crabtree Auditorium, Mount Allison University. Check our website for more details http://www.chebucto.ns.ca/Environment/ASFWB/ there have been others. Gerry Parker reported several cases of tourists being nipped as they fed coyotes near Cheticamp. Parks Canada has not released information on all incidents of coyote attacks. Now more people are noticing coyotes in strange places and they are feeling wary. In Bedford NS, coyotes were spotted on school property and parents were warned via a newsletter to keep their children safe. In Port Hawkesbury Canadian Wildlife Service wildlife technicians, Randy Hicks and Andrew coyotes were spotted near a wooded area adjacent to Hicks walked away bruised and with a broken foot respectively from this Tamarac Education Centre on the morning of March 8, 4- seater Cessna 172 on March 07, 2010 after engine problems forced an 2010, leading a local parent to approach school officials. emergency landing at Argyle, about 10 km from Yarmouth, Nova Scotia . Such stories can get people on edge. Bob Bancroft told of They were conducting low altitude coastal bird surveys in good weather on a Sunday. Their 26 year old pilot was treated in Halifax for severe when a coyote persisted in charging him. A rather large head injuries. (Tina Comeau - Yarmouth Vanguard Photo) Cape Breton University student met with coyotes when walking home in the dark and was relieved to find a house nearby. -
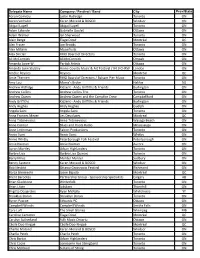
Aaron Comeau Justin Rutledge Toronto ON
Delegate Name Company / Festival / Band City Prov/State Aaron Comeau Justin Rutledge Toronto ON Aaron Verhulst Karen Morand & BOSCO Windsor ON Abigail Lapell Abigail Lapell Toronto ON Adam Lalonde Gabrielle Goulet Ottawa ON Adam Warner Ian Sherwood Toronto ON Alain Berge Élage Diouf Montréal QC Alec Fraser Jon Brooks Toronto ON Alex Millaire Moonfruits Ottawa ON Alex Sinclair FMO Board of Directors Toronto ON Ali McCormick Ali McCormick Ompah ON Amanda Lowe W. Partick Artists Ottawa ON Amanda Lynn Stubley Home County Music & Art Festival / 94.9 CHRW London ON Amélie Beyries Beyries Montréal QC Amie Therrien FMO Board of Directors / Balsam Pier Music Toronto ON Amy Lou Mama's Broke Halifax NS Andrew Aldridge Kidzent - Andy Griffiths & Friends Burlington ON Andrew Collins Andrew Collins Trio Toronto ON Andrew Queen Andrew Queen and the Campfire Crew Campbellford ON Andy Griffiths Kidzent - Andy Griffiths & Friends Burlington ON Andy Hughes Andy Hughes Guelph ON Angela Saini Angela Saini Toronto ON Anna Frances Meyer Les Deuxluxes Montreal QC Anna Tribinevicius Anna Tribinevicius Wasaga Beach ON Anne Connor Blues and Roots Radio Mississauga ON Anne Lederman Falcon Productions Toronto ON Annie Sumi Annie Sumi Whitby ON Annie Whitty Peterborough Folk Festival Peterborough ON Arnie Naiman Arnie Naiman Aurora ON Ayron Mortley Urban Highlanders Toronto ON Barbra Lica Barbra Lica Quintet Toronto ON Barry Miles Murder Murder Sudbury ON Benny Santoro Karen Morand & BOSCO Windsor ON Bob Nesbitt Ottawa Grassroots Festival Richmond ON Borza Ghomeshi -

Wildlife Hysteria: Nova Scotia's War on Coyotes
Wildlife Hysteria: Nova Scotia’s War on Coyotes By Billy MacDonald, Redtail Nature Awareness and David Orton, Green Web In Nova Scotia, coyotes are designated “other harvestable wildlife” and can be shot or otherwise killed year-round with no “bag limit”. There is also an NDP government-initiated subsidized trapping program, through a “pelt-incentive” of twenty dollars per dead coyote, for licenced trappers. We are informed that coyotes seen near communities, for example schools, “are to be captured and killed.” A Department of Natural Resources press release of Jan. 21, 2011, states that “More than 800 coyote pelts have been shipped to market this season, a 51 per cent increase over the same period last season.” Government media releases have spoken of aiming to kill 4,000 coyote! We are two people living at different, relatively isolated, rural locations in Pictou County, in Nova Scotia. Each of us has lived with coyotes – really wild dogs –in our immediate neighbourhoods, for over twenty- five years. We oppose the coyote fear-mongering and hatred in Nova Scotia, which encourages a dread of being in the woods where coyotes could roam. One of us has had hundreds of youth sleeping in woods at summer camps, with coyotes in the vicinity and with no incidents, for the past twenty years. Wildlife is “wild” and humans need to adapt to this. A measure of a supposedly civilized society should be human tolerance and co-existence with all other species, and concern for their well-being, not just for humans and their domesticated pets. We need a deeper ecological awareness. -

Big-Name Musicians Croon for Clean
ART ENT GULF ISLANDS DRIFTWOOD .o. WEDNESDAY, SEPTEMBER 22,2004 .o. PAGE 81 Big-name musicians croon for clean air By MITCHELL SHERRIN improvisations it sounded Bachman's You Ain't Seen Staff Writer like he was playing two gui Nothing Yet, he joined in the Islanders counted heav tars. And lyrics from songs fun on stage and stuttered ily among a keen crowd like Don't Let it Bring You along with Stewart as a high of 2,300 who roared with Down seemed to take on a light to the evening. delight to see rock legends new weight in light of the · BNL wrapped up the Neil Young, Randy Bach environmental cause behind four-and-a-half hour concert man, Ta1 Bachman and the the benefit: "It's only castles with a high-energy perfor Barenaked Ladies perform burning, find someone who's mance highlighted by clever at the Clean Air Concert in turning and you will come puns, comedic raps, jazzy Duncan Friday. around." harmonies and bass-heavy Headlining the show at Young even called his wife grooves. the Cowichan Centre Arena, Pegi to join him with vocals · Their set included lots of Young appeared on a Van on the classics Hmnan High pop favourites like Hello couver Island stage for the way, Old King and Goin' City, One Week, Blame it first time when he threw in Back. on Me, Some Fantastic and his hat to help environmen After a thunderous stand Alternative Girlfriend. talists battle the nearby Crof ing ovation, Young returned The boys from BNL even ton pulp mill. -

Should You Fear the Pizzly Bear? - Nytimes.Com
Should You Fear the Pizzly Bear? - NYTimes.com http://www.nytimes.com/2014/08/17/magazine/should-you-fear-the-pizz... In New England today, trees cover more land than they have at any time since the colonial era. Roughly 80 percent of the region is now forested, compared with just 30 percent in the late 19th century. Moose and turkey again roam the backwoods. Beavers, long ago driven from the area by trappers seeking pelts, once more dam streams. White-tailed deer are so numerous that they are often considered pests. And an unlikely predator has crept back into the woods, too: what some have called the coywolf. It is both old and new — roughly one-quarter wolf and two-thirds coyote, with the rest being dog. The animal comes from an area above the Great Lakes, where wolves and coyotes live — and sometimes breed — together. At one end of this canid continuum, there are wolves with coyote genes in their makeup; at the other, there are coyotes with wolf genes. Another source of genetic ingredients comes from farther north, where the gray wolf, a migrant species originally from Eurasia, resides. “We call it canis soup,” says Bradley White, a scientist at Trent University in Peterborough, Ontario, referring to the wolf-coyote hybrid population. The creation story White and his colleagues have pieced together begins during European colonization, when the Eastern wolf was hunted and poisoned out of existence in its native Northeast. A remnant population — “loyalists” is how White refers to them — migrated to Canada. At the same time, coyotes, native to the Great Plains, began pushing eastward and mated with the refugee wolves. -

Conference Guide
BOARD OF DIRECTORS 2018/19 EXECUTIVE COMMITTEE DIRECTORS CO-PRESIDENT Tim Des [email protected] Amie [email protected] Richard [email protected] CO-PRESIDENT Liz Scott.............................................lizscott555@gmail.com Preetam [email protected] Syma [email protected] VICE PRESIDENT Rosalyn [email protected] PAST PRESIDENTS TREASURER Paul [email protected] Katherine Partridge Carolyn Bigley Rachel Barreca Bill Marshall SECRETARY Max Merrifield............................................director@nlfb.ca Alex Sinclair Magoo Scott Merrifield Jim McMillan MEMBER-AT-LARGE Paul Mills Emma Jane [email protected] Aengus Finnan STAFF Sam Baijal Doug McArthur EXECUTIVE DIRECTOR Alka [email protected] Warren Robinson OFFICE ADMINISTRATOR Lianne Rancourt from May - August PAST EXECUTIVE DIRECTORS Kyle Young from September - October Peter MacDonald Erin Benjamin FOLK NORTH COORDINATOR Rachel [email protected] VOLUNTEER COORDINATOR PAST ESTELLE KLEIN AWARD RECIPIENTS Catherine [email protected] Bill Garrett Stan Rogers DEVELOPING ARTISTS PROGRAM COORDINATOR Magoo Richard Flohil Treasa [email protected] -

Page 1 of 1 CTV.Ca
CTV.ca - Saskatchewan minister defends coyote bounty - CTV News, Shows and Sports -- Canadian T... Page 1 of 1 Saskatchewan minister defends coyote bounty The Canadian Press Updated: Thu. May. 27 2010 6:50 AM ET REGINA — More than 71,000 coyotes that had a bounty on their heads have been killed in Saskatchewan, but critics say the number is appalling and doesn't solve a predator problem. Agriculture Minister Bob Bjornerud said Wednesday that the number of coyotes destroyed under the pilot program was a surprise. But he was pleased with the results and said the cull was necessary because the animals had become a threat to livestock and farm families. "We certainly didn't start out to eradicate coyotes in the province. That wasn't our intention at all," said Bjornerud. "But we had to do something with the number of problems that it was creating with ... calves and sheep and things like that across the province. "The other problem was that there was almost no respect in the coyote population. They were packing up, coming right into yards. We talked before about them coming onto porches and eating out of the dog dish and in some cases killing the pets. The farm dog ... was being lured out by packs and being killed." The Saskatchewan government introduced its five-month coyote control program last November. It offered hunters, farmers and ranchers $20 per dead coyote as long as all four paws were brought in. The final cost to the province will be about $1.5 million. Bjornerud said the next step is to gauge the cull's effectiveness, but that's difficult because no one knows how many coyotes are in Saskatchewan. -

147 June to August 2012
THE HALIFAX FIELD NATURALIST No. 147 June to August 2012 In This Issue ...................................................... 2 HFN Field Trips ............................................... 6 Editorial & Nature Notes ................................ 3 Almanac ............................................................ 9 HFN Talks ......................................................... 4 Hfx Tide Table: July to September ...............11 Return address: HFN, c/o NS Museum of Natural History, 1747 Summer Street, Halifax, NS, B3H 3A6 1 Summer 2012, #147 is incorporated under the Nova Scotia HFN ADDRESS Societies Act and holds Registered Halifax Field Naturalists, c/o N.S. Museum of Natural History, Charity status with Canada Revenue 1747 Summer St., Hfx, N.S., B3H 3A6 Email: [email protected] HFN Website: halifaxfieldnaturalists.ca Agency. Tax-creditable receipts will be issued for individual and corporate gifts. HFN is an affiliate of Nature Canada and NNS ADDRESS an organisational member of Nature Nova Scotia, the provin- Nature Nova Scotia, c/o N.S. Museum of Natural History, 1747 cial umbrella association for naturalist groups in Nova Sco- Summer St., Halifax, N.S., B3H 3A6 Email: [email protected] (Doug Linzey, NNS Secretary and tia. Objectives are to encourage a greater appreciation and Newsletter Editor) Website: naturens.ca understanding of Nova Scotia’s natural history, both within the EXECUTIVE 2011/2012 membership of HFN and in the public at large, and to represent President Janet Dalton .......................................443-7617 the interests of naturalists by encouraging the conservation of Vice-President Clarence Stevens ...............................864-0802 Nova Scotia’s natural resources. Meetings are held, except for Treasurer Doris Balch .........................................423-3342 July and August, on the first Thursday of every month at 7:30 Secretary Richard Beazley .................................429-6626 p.m. -

16263 Wff2011annualreportcover Finpths.Indd
TABLE OF CONTENTS 2010 CHAIR’S MESSAGE 2 2010 EXECUTIVE DIRECTOR’S MESSAGE 3 2010-11 SNAPSHOT 4 2010-11 BOARD OF DIRECTORS 5 I. WINNIPEG FOLK FESTIVAL ORGANIZATIONAL PROFILE 6 II. FESTIVAL HISTORY AND ACTIVITIES 8 III. TEN-YEAR FINANCIAL HISTORY 10 IV. 2010-11 FINANCIAL OVERVIEW 11 V. 2010 WINNIPEG FOLK FESTIVAL 12 HISTORY OF PAID ATTENDANCE 21 2010 WINNIPEG FOLK FESTIVAL RECOGNITION AWARDS 22 VI. YEAR-ROUND ACTIVITIES 23 VII. WINNIPEG FOLK FESTIVAL MUSIC STORE 27 VIII. RESOURCE DEVELOPMENT 28 WINNIPEG FOLK FESTIVAL 2010 SPONSORS 30 IX. STRATEGIC INITIATIVES 32 WINNIPEG FOLK FESTIVAL 2010-11 STAFF 34 PAST PERFORMERS 1974-2010 35 APPENDIX 49 RESPONSIBILITY FOR FINANCIAL STATEMENTS 52 2010 Chair’s Message 10 Our festival started as a simple one-time event in 1974, and 37 years later, it still remains as the centre of our universe, constantly evolving, changing and growing over the years to meet our changing audiences, while staying true to the values that best represent our festival. Much to our surprise (and pleasure), we seem to be garnering all kinds of national and international recognition in the form of Top 10 lists and musical and tourism awards for simply doing the things that we love to do. Our success rests in not being comfortable with what we did yesterday, but what we might be today and tomorrow. The 2010 event was spectacular again in the ways we have always expected: Five days of beautiful weather; a wonderful harmony between our audience, 2,500 volunteers, and artists; a wonderful musical experience and the ongoing joys of discovery offered up yet again by Chris Frayer, our Artistic Director guru; another record-breaking crowd that pushed us almost to site capacity; and another really successful year financially.