The Long-Term Distribution, Habitat Use and Site Fidelity of Bottlenose
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
Scottish Samurai Trail the Story of Thomas Blake Glover
Scottish Samurai Trail The story of Thomas Blake Glover #aberdeentrails Model of Jho Sho Maru #aberdeentrails Much has been written about the life and times of Thomas Blake Glover, and many myths have grown up around him. This guide has been produced to introduce his story and to inspire you to learn more about the era, the man, and some of places associated with him. Thomas’s links with Japan, and the changes that country went through in the latter half 19th century are rightly celebrated there, and his home in Nagasaki is Above: Thomas Blake Glover wearing the Order of the Rising Sun preserved as a museum, in extensive parkland known as Glover Garden. Courtesy Nagasaki Museum of History and Culture Both Aberdeen and Fraserburgh have connections to Glover and this trail guide covers both locations. Enjoy discovering about Thomas, his early life Cover: Thomas Blake Glover Courtesy Glover Garden in Scotland, and finding out about our connections to Japan! Accessibility This trail is accessible but has occasional steep parts / uneven ground. Picture Credits Transport All images © Aberdeen City Council unless otherwise stated. The historical images in the first section are courtesy First Bus 20 runs through Old Aberdeen to Don Street near Brig O’ Balgownie. Nagasaki Museum of History and Culture First Bus 15 runs to Footdee, returning by York Street. First Bus 1 & 2 run to Bridge of Don via King Street and Ellon Road. Images at 3 & 4: Courtesy of Aberdeen City Libraries/Silver City Vault All services go to/from Union Street. www.silvercityvault.org.uk Stagecoach 67/68 runs between Fraserburgh and Aberdeen Bus Station via King Street and Ellon Road. -

ADDRESSING CONCERNS RAISED by RSPB SCOTLAND .88 27 Addressing Concerns Raised by RSPB Scotland
Annex B - Appropriate Assessment – Moray West Offshore Wind Farm T: +44 (0)300 244 5046 E: [email protected] SCOTTISH MINISTERS ASSESSMENT OF THE PROJECT’S IMPLICATIONS FOR DESIGNATED SPECIAL AREAS OF CONSERVATION (“SAC”), SPECIAL PROTECTION AREAS (“SPA”) AND PROPOSED SPECIAL PROTECTION AREAS (“pSPA”) IN VIEW OF THE SITES’ CONSERVATION OBJECTIVES APPLICATION FOR CONSENT UNDER SECTION 36 OF THE ELECTRICITY ACT 1989 (AS AMENDED) AND FOR MARINE LICENCES UNDER THE MARINE (SCOTLAND) ACT 2010 AND MARINE AND COASTAL ACCESS ACT 2009 FOR THE CONSTRUCTION AND OPERATION OF THE MORAY WEST OFFSHORE WIND FARM AND ASSOCIATED OFFSHORE TRANSMISSION INFRASTRUCTURE SITE DETAILS: MORAY WEST OFFSHORE WIND FARM AND EXPORT CABLE CORRIDOR BOUNDARY – APPROXIMATELY 22.5KM EAST OF THE CAITHNESS COASTLINE IN THE OUTER MORAY FIRTH Name Assessor or Approver Date Fiona Mackintosh Assessor 15 April 2019 Ross Culloch Assessor 15 April 2019 Tom Evans Assessor 15 April 2019 Gayle Holland Approver 26 April 2019 Annex B - Appropriate Assessment – Moray West Offshore Wind Farm TABLE OF CONTENTS SECTION 1: BACKGROUND ..................................................................................... 2 1 Introduction ................................................................................................................ 2 2 Appropriate assessment (“AA”) conclusion ................................................................ 2 3 Background to including assessment of proposed SPAs ........................................... 3 4 Details of proposed operation -

List of Consultees and Issues.Xlsx
Name / Organisation Issue Mr Ian Adams Climate change Policy C1 Using resources in buildings Mr Ian Adams Shaping Formartine Newburgh Mr Iain Adams Natural Heritage and Landscape Policy E2 Landscape Mr Ian Adams Shaping Formartine Newburgh Mr Michael Adams Natural Heritage and Landscape Policy E2 Landscape Ms Melissa Adams Shaping Marr Banchory Ms Faye‐Marie Adams Shaping Garioch Blackburn Mr Iain Adams Shaping Marr Banchory Michael Adams Natural Heritage and Landscape Policy E2 Landscape Ms Melissa Adams Natural Heritage and Landscape Policy E2 Landscape Mr Michael Adams Shaping Marr Banchory Mr John Agnew Shaping Kincardine and Mearns Stonehaven Mr John Agnew Shaping Kincardine and Mearns Stonehaven Ms Ruth Allan Shaping Banff and Buchan Cairnbulg and Inverallochy Ruth Allan Shaping Banff and Buchan Cairnbulg and Inverallochy Mrs Susannah Almeida Shaping Banff and Buchan Banff Ms Linda Alves Shaping Buchan Hatton Mrs Michelle Anderson Shaping Kincardine and Mearns Luthermuir Mr Murdoch Anderson Shaping Kincardine and Mearns Luthermuir Mrs Janette Anderson Shaping Kincardine and Mearns Luthermuir Miss Hazel Anderson Shaping Kincardine and Mearns Luthermuir J Angus Shaping Banff and Buchan Cairnbulg and Inverallochy Mrs Eeva‐Kaisa Arter Shaping Kincardine and Mearns Mill of Uras Mrs Eeva‐Kaisa Arter Shaping Kincardine and Mearns Mill of Uras Mr Robert Bain Shaping Garioch Kemnay K Baird Shaping Banff and Buchan Cairnbulg and Inverallochy Rachel Banks Shaping Formartine Balmedie Mrs Valerie Banks Shaping Formartine Balmedie Valerie Banks -

The Dalradian Rocks of the North-East Grampian Highlands of Scotland
Revised Manuscript 8/7/12 Click here to view linked References 1 2 3 4 5 The Dalradian rocks of the north-east Grampian 6 7 Highlands of Scotland 8 9 D. Stephenson, J.R. Mendum, D.J. Fettes, C.G. Smith, D. Gould, 10 11 P.W.G. Tanner and R.A. Smith 12 13 * David Stephenson British Geological Survey, Murchison House, 14 West Mains Road, Edinburgh EH9 3LA. 15 [email protected] 16 0131 650 0323 17 John R. Mendum British Geological Survey, Murchison House, West 18 Mains Road, Edinburgh EH9 3LA. 19 Douglas J. Fettes British Geological Survey, Murchison House, West 20 Mains Road, Edinburgh EH9 3LA. 21 C. Graham Smith Border Geo-Science, 1 Caplaw Way, Penicuik, 22 Midlothian EH26 9JE; formerly British Geological Survey, Edinburgh. 23 David Gould formerly British Geological Survey, Edinburgh. 24 P.W. Geoff Tanner Department of Geographical and Earth Sciences, 25 University of Glasgow, Gregory Building, Lilybank Gardens, Glasgow 26 27 G12 8QQ. 28 Richard A. Smith formerly British Geological Survey, Edinburgh. 29 30 * Corresponding author 31 32 Keywords: 33 Geological Conservation Review 34 North-east Grampian Highlands 35 Dalradian Supergroup 36 Lithostratigraphy 37 Structural geology 38 Metamorphism 39 40 41 ABSTRACT 42 43 The North-east Grampian Highlands, as described here, are bounded 44 to the north-west by the Grampian Group outcrop of the Northern 45 Grampian Highlands and to the south by the Southern Highland Group 46 outcrop in the Highland Border region. The Dalradian succession 47 therefore encompasses the whole of the Appin and Argyll groups, but 48 also includes an extensive outlier of Southern Highland Group 49 strata in the north of the region. -

Pre-Application Consultation (PAC) Report
Moray West Offshore Wind Farm and Offshore Transmission Infrastructure (OfTI) Pre-application Consultation (PAC) Report This Pre-application Consultation (PAC) Report has been prepared in accordance with requirements of the Marine Licencing (Pre-application Consultation) (Scotland) Regulations 2013 in support of an application for a Marine Licence for the Moray West Offshore Transmission Infrastructure (OfTI) components of the Moray West Offshore Wind Farm. 1. Proposed Licensable Marine Activity Please describe below or, where there is insufficient space, in a document attached to this form the proposed licensable marine activity, including its location Moray Offshore Windfarm (West) Limited (known as Moray West) are proposing to develop the following licensable marine activities/project; Moray West Offshore Wind Farm and Offshore Transmission Infrastructure (OfTI). Maps showing the location of the Offshore Wind Farm and OfTI are presented in Appendix A. The Moray West Offshore Wind Farm will comprise up to 85 Wind Turbine Generators (WTGs), associated substructures and seabed foundations, inter-array cables and any scour protection around substructures or cable protection. The OfTI comprises up to two Offshore Substation Platforms (OSPs) which will be located within the Moray West Site, OSP interconnector cables and two offshore export cable circuits which will be located within the Offshore Export Cable Corridor and will be used to transmit the electricity generated by the offshore wind farm to shore. The offshore export cable circuits come ashore in the Landfall Area which is located on the Aberdeenshire Coast between Findlater Castle and Redhythe Point, approximately 65 km south of the Moray West Site. The Marine Licensing (Pre-application Consultation) (Scotland) Regulations 2013 apply to all licensable marine activities (as defined under the Marine (Scotland) Act 2010) within Scottish marine area, the area of sea within the seaward limits of the territorial sea (0 – 12 nautical miles [nm]). -

Banff Castle
UE 12 2010 - ISS insideinside thisthis issueissue .. .. .. newsnews fromfrom aroundaround thethe areaarea .. .. .. TransportTransport newsnews .. .. .. andand lotslots moremore partnershipupdate Chairman’s Letter Design: Kay Beaton, elcome to the latest edition of the Banffshire Partnership PURPLEcreativedesign WNewsletter. During the year one of the longest serving directors Eddie Bruce Printed by had to stand down due to ill-health. Eddie was a very valued Halcon, Aberdeen member of the board and I would like to record our thanks for community transport Paper his contribution and wise counsel over the years. Printed on environ- Also during the year Evelyn Elphinstone, our administrator and book-keeper retired. Evelyn had worked for the Partnership for many mentally friendly paper. years and I would also like to record our thanks for her hard work and Woodpulp sourced from dedication over the years. sustainable forests. This has been a busy and challenging year for BPL. Once again we entered into a formal Service Level Agreement with Aberdeenshire Council which commenced on 1st April and Board Of Directors runs to 31st March 2010. This is core funding for the Partnership which allows it to carry Directors can be out its very important tasks helping many community groups throughout our operational “keeping the community moving” area. contacted through the With the expected squeeze on local government finances in the coming years there Partnership is no guarantee that such funding will endure at the required level. However, both community use minibus office - 01261 843286. Aberdeenshire Council and the Local Rural Partnerships across the shire are keen to ensure that they survive any reduced funding from the Scottish Government. -

Human Environment Baseline.Pdf
Moray Offshore Renewables Limited - Environmental Statement Telford, Stevenson and MacColl Offshore Wind Farms and Transmission Infrastructure 5 Human Environment 5.1 Commercial Fisheries 5.1 5.1.1 Introduction 5.1.1.1 This chapter summarises the baseline study of commercial fishing activities, including salmon and sea trout fisheries, in the vicinity of the three proposed development sites (Telford, Stevenson and MacColl) and the offshore transmission infrastructure (OfTI). For the purpose of this study, commercial fishing is defined as CHAPTER any legal fishing activity undertaken for declared taxable profit. 5.1.1.2 The following technical appendices support this chapter and can be found as: Technical Appendix 4.3 B (Salmon and Sea Trout Ecology Technical Report). Technical Appendix 5.1 A (Commercial Fisheries Technical Report). 5.1.1.3 For the purposes of this assessment, salmon and sea trout fisheries in the Moray Firth are separately addressed to other commercial fisheries, as a result of their being located largely in-river (with the exception of some coastal netting) and being different in nature to the majority of marine commercial fishing activities. In addition, due to the migratory behaviour of salmon and sea trout, fisheries have been assessed for all rivers flowing into the Moray Firth. It is also recognised that salmon is a qualifying feature or primary reason for Special Area of Conservation (SAC) site selection of the following rivers in the Moray Firth: Berriedale and Langwell Waters SAC (primary reason); River Moriston -
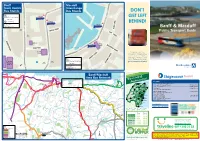
Banff and Macduff Public Transport Guide October 2015
Banff_Town_Centre_Map.ai 1 16/10/2015 11:44 Macduff_Town_Centre_Map.ai 1 16/10/2015 11:43 Banff BOYNDIE STREET Macduff Town©P1ndar Centre ©P1ndar © Interchange©P1ndar ©P1ndar © Bus Stands Bus Stands REET DON’T CARMELITE STREET T S S Key A S E Low Street ©P1ndar Road served by bus B9142 LOW STREE GET LEFT O’N A98 HIGH STREET Bus stop K B E Car parking ©P1ndar Low Street N CROO BEHIND! A Don’t get left behind Contains Ordnance Survey data LA Nicols Brae ©P1ndar © Crown copyright 2015 L i Digital Cartography by Pindar Creative www.pindarcreative.co.uk©P1ndar ©P1ndar © ©P1ndar ©P1ndar © HOO Banff & Macduff SC High T Street ©P1ndar BRIDGE STREET DUF Public Transport Guide F 8 S 9 TREET High A ©P1ndar Street October 2015 WALKER AVENUE Library ©P1ndar B Town Town Hall I ©P1ndar ©P1ndar © ©P1ndar ©P1ndarHall NST © IT UT ©P1ndar I To receive advanced notification of changes to BACK PATH Hutcheon Street ON STREET S bus services in Aberdeenshire by email, E TR sign up for our free alert service at OR EE Court SH T www.aberdeenshire.gov.uk/publictransport/status/ EET To receive advanced House R T We are currently in the process of ET notificationequipping of all buschanges stops in to bus S Aberdeenshire with QR Codes and NFC Technology. TRE services in Aberdeenshire by W This will allow you to look up bus S times from your stop for free* in O seconds using your Smartphone. L L email, sign up for our free alert CHURCH STREET L Look for symbols like these Key I service at www.aberdeenshire.at the bus stop Just scan the top QR Code, or if you Airlie Road served by bus LLYM have a smartphone equipped with NFC gov.uk/publictransport/status/technology, hold it against the indicated Banff ©P1ndar E 9142 Gardens Airlie G area to take you to a page showing B the departure times from your stop. -

Housing Development Opportunities
Housing Development Opportunities Seafield Estate Zoned ResidentialSites For Sale From 0.24 - 17.93 hectares Sites available in: • Fordyce • Cullen • Portknockie • Portsoy • Findochty • Sandend • Buckie • Whitehills • Rathven Introduction As part of this process Seafield and Strathspey Estates are working with Cushman & Wakefield to promote owned sites and offer information Seafield and Strathspey Estates, who have their to interested parties. The purpose of this document Estate office at Cullen in Banffshire, are one of is to illustrate the scale and diversity of opportunities Scotland’s largest and leading agricultural in North East Scotland. providers managing an operation extending over some 85,000 acres of Scotland’s finest scenery Provided for each site (designated as housing diversified into agricultural, woodland and tour- land in the relevant Local Plan) is the preliminary ism/leisure related uses. information that you may require. Cushman & Wakefield can assist with additional information, As part of an ongoing strategy the Estates have if available, but the basis of this document is to worked with Local Councils to bring forward land encourage an active response leading to development suitable for development, enhancing opportunity. opportunities for inward investment and offering local businesses land for development. CONTENTS Cullen................................................1 Portsoy........................................2 Sandend.........................................3 Whitehills.................4 Fordyce.............................5 -

Open Space Assessment Sheet Community Council: Aberchirder-Marnoch Settlement: Aberchirder
Open Space Assessment Sheet Community Council: Aberchirder-Marnoch Settlement: Aberchirder Access Type Site Management/ Level Of Opportunities Or Site Score Site Name Site Description (Full, Relative Customer Type Problems Ownership (If Known) Usage Recommendations (Out Of 100) Or Restricted) Very large green Anderson Aberdeenshire park with play Full Public Children. Parents. Often. Goal Lack of play Wider variety of 62 Mclenan Park Council area and football Access Local community areas worn facilities facilities pitch Green space Residents of Walker Court Aberdeenshire Full Public within housing surrounding Unclear None More planting 57 Amenity Area Council Access area houses Causeway Aberdeenshire Small play area Full Public Unclear. New Lack of play 50 Local children None End Council among houses Access play area facilities Poorest Play area within Residents of Softer surface Hardie Court Full Public Often. Play 51 Unknown residential surrounding None around play Play Area Access area worn Best housing estate houses equipment Parkview Aberdeenshire Full Public Lack of play 74 Playing field Local community Often None Playing Field Council Access facilities Poorest Aberchirder Aberchirder Bowling Restricted Members and Well used 80 Bowling green None More planting Bowling Green Club Access visitors of club during season Best Unclear Saint Marnan Full Public No Disabled Episcopal Church Churchyard Parishioners During Site Planting 56 Church Access Access Visit Open Space Assessment Sheet Community Council: Aberdour and Tyrie Settlement: -

BPL-Partnership-Update-14
ISSUE 14 - 2012 PARTNERSHIP UPDATE WWW.BANFFSHIREPARTNERS.CO.UK INSIDE THIS ISSUE . NEWS FROM AROUND THE FREEAREA . TRANSPORT NEWS . AND LOTS MORE PARTNERSHIP BANFFSHIRE PARTNERSHIP LIMITED UPDATE Banffshire Partnership Ltd. Chairman’s Letter The Old School, Boyndie, Welcome to the new edition to make the operation more Banff, AB45 2JT T & F: 01261 843598 of the Banffshire Partnership cost-effective. This is even developing communities E: [email protected] Yearbook. As expected, it more important now as we has been a very challenging have seen a reduction in our BOARD OF DIRECTORS year for everybody at the budget allocation for this year Directors can be contacted Partnership and I would and expect further reductions through the Partnership office - Partnership and it has assisted 01261 843598. like to thank Duncan Leece, in the coming years. Directors meet bi-monthly his staff and especially the many community groups with (or more often if necessary). volunteers for all their efforts TRAINING projects, business plans and “support for an active community” in meeting those challenges. For many years we have funding applications over the CHAIR Robin Maddock provided transport-related years. SECRETARY Iain Taylor TREASURER Ross Balharry COMMUNITY TRANSPORT training to minibus drivers We have a paid contract community development Denis Askham Since the Community Transport and Passenger Assistants. This Sandy Duncan side of the business started year, in an effort to broaden with Aberdeenshire Council Roger Goodyear in 1999 it has grown steadily our range, we are now able to provide this resource and marketing support Ian Gray without that income we would Anne McArthur and is now the main activity to offer a range of training of Banffshire Partnership. -

The Perfect Rural Idyll with Period Family House, Extensive Versatile
THE PERFECT RURAL IDYLL WITH PERIOD FAMILY HOUSE, EXTENSIVE VERSATILE BUILDINGS AND LAND lintmill of boyne, banff, ab45 2tl THE PERFECT RURAL IDYLL WITH PERIOD FAMILY HOUSE, EXTENSIVE VERSATILE BUILDINGS AND LAND lintmill of boyne, banff, ab45 2tl Summary Lintmill of Boyne House 2 reception rooms u 4 bedrooms u 3 bathrooms Outbuildings u workshop and gym Steading Traditional stone and slate u 6 stables u tack room u feed room u store u workshop u barn u music room and loft u cart sheds/stores Land 4 acre paddock u sand school Approximately 6.5 acres in total for sale as a whole EPC = E Portsoy 2 miles, Banff 6 miles, Aberdeen 50 miles, Inverness 65 miles Location Lintmill of Boyne is situated in a peaceful and secluded rural location about a mile inland from the coastline of the Moray Firth and about 2 miles to the east of the village of Portsoy. The local area is famed for its breathtaking scenery: there are several picturesque villages situated along the rugged Moray/Banffshire coastline which is interspersed with cliffs and sheltered sandy beaches. There is a blue flag beach at Cullen and further spectacular sheltered coves at Sandend, Banff and Pennan. The nearby village of Portsoy provides basic everyday facilities including a supermarket and primary school. The attractive and historic harbour is home to the Scottish Traditional Boat Festival every year which engages up to 16,000 people in the celebrations. To the east is the nearby historic town of Banff which lies at the mouth of the River Deveron.