PTH Promotes Rabbit Tibial Fracture Healing Via the Notch Signaling Pathway Q.-H
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

FCC RF EXPOSURE REPORT for CONSUMER CAMERA
FCC RF EXPOSURE REPORT For CONSUMER CAMERA MODEL NUMBER: DH-IPC-D26P, IPC-D26P, DH-IPC-D26N, IPC-D26N, TD1, TD1C, DH-IPC-D26, IPC-D26 PROJECT NUMBER: 4788111562 REPORT NUMBER: 4788111562-7 FCC ID: SVNDH-IPC-DX6 ISSUE DATE: Dec. 7, 2017 Prepared for Zhejiang Dahua Vision Technology Co., Ltd. Prepared by UL Verification Services (Guangzhou) Co., Ltd, Song Shan Lake Branch Room 101, Building 10, Innovation Technology Park, Song Shan Lake Hi tech Development Zone, Dongguan, 523808, China Tel: +86 769 33817100 Fax: +86 769 33244054 Website: www.ul.com The results reported herein have been performed in accordance with the laboratory’s terms of accreditation. This report shall not be reproduced except in full without the written approval of the Laboratory. The results in this report apply to the test sample(s) mentioned above at the time of the testing period only and are not to be used to indicate applicability to other similar products. This report does not imply that the product(s) has met the criteria for certification. REPORT NO: 4788111562-7 DATE: Dec. 7, 2017 FCC ID: SVNDH-IPC-DX6 TABLE OF CONTENTS 1. ATTESTATION OF TEST RESULTS ................................................................................. 3 2. TEST METHODOLOGY ..................................................................................................... 4 3. FACILITIES AND ACCREDITATION ................................................................................. 4 4. REQUIREMENT ................................................................................................................. 5 Page 2 of 6 UL Verification Services (Guangzhou) Co., Ltd, Song Shan Lake Branch This report shall not be reproduced except in full, without the written approval of UL Verification Services (Guangzhou) Co., Ltd, Song Shan Lake Branch. REPORT NO: 4788111562-7 DATE: Dec. 7, 2017 FCC ID: SVNDH-IPC-DX6 1. ATTESTATION OF TEST RESULTS Applicant Information Company Name: Zhejiang Dahua Vision Technology Co., Ltd. -

Luxembourg - 4Th -9Th August 2019
Abstract proposal for 2017 IGU Urban Commission Meeting Luxembourg - 4th -9th August 2019 Abstract proposal for 2019 IGU Urban Commission Meeting Luxembourg – 4th- 9th August 2019 2019 IGU Urban Commission Annual Meeting Urban Challenges in a Complex WorlD The urban geographies of the new economy, services inDustries, anD financial marketplaces AUTHORS: Lu Xu, Weibin Peng Paper Title: The Effect of Spatial Diversity on the New Properties Price in Hangzhou under the Housing Market Regulation -An Empirical Analysis Based on POI Data Session number: 7 Urban Governance, planning and participative democracy ExtenDeD abstract Abstract: Facing soaring housing prices, a new return of strict market regulatory measures has been implemented by the Chinese government since 2018. The municipal government of Hangzhou also released a lottery policy on new properties market armed to curb its surging housing prices. Based on the POI data in Hangzhou, this paper uses the count model to obtain the number of POI distribution points within 2 km scope of different lottery properties in the metropolitan areas since 4th April 2018,proposes three methods using OLS, GWR and Kriging interpolation method to empirically study the spatial influential factors on the new properties’ prices under the administrative interventions as well as purchase restrictions. Keywords: Spatial diversity, Price regulation, the lottery of new properties; POI Introduction Since the recovery of the Chinese real estate market in 2016, the housing prices in Hangzhou which as the capital of Zhejiang Province and the city hosted the G20 summit has been Soaring. The serial surging housing prices and rising serious household indebtedness are the vehicles through which macroeconomic policy works?. -
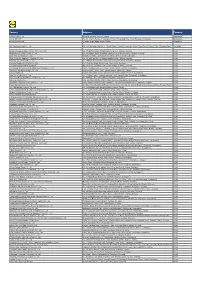
Factory Address Country
Factory Address Country Durable Plastic Ltd. Mulgaon, Kaligonj, Gazipur, Dhaka Bangladesh Lhotse (BD) Ltd. Plot No. 60&61, Sector -3, Karnaphuli Export Processing Zone, North Potenga, Chittagong Bangladesh Bengal Plastics Ltd. Yearpur, Zirabo Bazar, Savar, Dhaka Bangladesh ASF Sporting Goods Co., Ltd. Km 38.5, National Road No. 3, Thlork Village, Chonrok Commune, Korng Pisey District, Konrrg Pisey, Kampong Speu Cambodia Ningbo Zhongyuan Alljoy Fishing Tackle Co., Ltd. No. 416 Binhai Road, Hangzhou Bay New Zone, Ningbo, Zhejiang China Ningbo Energy Power Tools Co., Ltd. No. 50 Dongbei Road, Dongqiao Industrial Zone, Haishu District, Ningbo, Zhejiang China Junhe Pumps Holding Co., Ltd. Wanzhong Villiage, Jishigang Town, Haishu District, Ningbo, Zhejiang China Skybest Electric Appliance (Suzhou) Co., Ltd. No. 18 Hua Hong Street, Suzhou Industrial Park, Suzhou, Jiangsu China Zhejiang Safun Industrial Co., Ltd. No. 7 Mingyuannan Road, Economic Development Zone, Yongkang, Zhejiang China Zhejiang Dingxin Arts&Crafts Co., Ltd. No. 21 Linxian Road, Baishuiyang Town, Linhai, Zhejiang China Zhejiang Natural Outdoor Goods Inc. Xiacao Village, Pingqiao Town, Tiantai County, Taizhou, Zhejiang China Guangdong Xinbao Electrical Appliances Holdings Co., Ltd. South Zhenghe Road, Leliu Town, Shunde District, Foshan, Guangdong China Yangzhou Juli Sports Articles Co., Ltd. Fudong Village, Xiaoji Town, Jiangdu District, Yangzhou, Jiangsu China Eyarn Lighting Ltd. Yaying Gang, Shixi Village, Shishan Town, Nanhai District, Foshan, Guangdong China Lipan Gift & Lighting Co., Ltd. No. 2 Guliao Road 3, Science Industrial Zone, Tangxia Town, Dongguan, Guangdong China Zhan Jiang Kang Nian Rubber Product Co., Ltd. No. 85 Middle Shen Chuan Road, Zhanjiang, Guangdong China Ansen Electronics Co. Ning Tau Administrative District, Qiao Tau Zhen, Dongguan, Guangdong China Changshu Tongrun Auto Accessory Co., Ltd. -

Spatial Distribution Pattern of Minshuku in the Urban Agglomeration of Yangtze River Delta
The Frontiers of Society, Science and Technology ISSN 2616-7433 Vol. 3, Issue 1: 23-35, DOI: 10.25236/FSST.2021.030106 Spatial Distribution Pattern of Minshuku in the Urban Agglomeration of Yangtze River Delta Yuxin Chen, Yuegang Chen Shanghai University, Shanghai 200444, China Abstract: The city cluster in Yangtze River Delta is the core area of China's modernization and economic development. The industry of Bed and Breakfast (B&B) in this area is relatively developed, and the distribution and spatial pattern of Minshuku will also get much attention. Earlier literature tried more to explore the influence of individual characteristics of Minshuku (such as the design style of Minshuku, etc.) on Minshuku. However, the development of Minshuku has a cluster effect, and the distribution of domestic B&Bs is very unbalanced. Analyzing the differences in the distribution of Minshuku and their causes can help the development of the backward areas and maintain the advantages of the developed areas in the industry of Minshuku. This article finds that the distribution of Minshuku is clustered in certain areas by presenting the overall spatial distribution of Minshuku and cultural attractions in Yangtze River Delta and the respective distribution of 27 cities. For example, Minshuku in the central and eastern parts of Yangtze River Delta are more concentrated, so are the scenic spots in these areas. There are also several concentrated Minshuku areas in other parts of Yangtze River Delta, but the number is significantly less than that of the central and eastern regions. Keywords: Minshuku, Yangtze River Delta, Spatial distribution, Concentrated distribution 1. -

CERTIFICATE Certificat - Certificado- Сертификат - Zertifikat - 證書
ISET S.r.l. Unipersonale Sede Legale e Uffici Cap. soc. i.v. € 10.200,00 Via Donatori di sangue, 9 - 46024 Moglia (MN) Cod. Fisc. e P.IVA Reg. Imprese 02 332 750 369 Tel. e fax +39 (0)376 598963 REA 02 332 750 369 www.iset-italia.com [email protected] Cap. soc. i.v. MN 0221098 CERTIFICATE Certificat - Certificado- Сертификат - Zertifikat - 證書 1) APPLICANT: (who finally puts the product on the market) Hangzhou IECHO Science & Technology Co., Ltd No.1 Building, NO.1 Weiye Road, Binjiang District, Hangzhou city, Zhejiang Province, China. MANUFACTURER: Hangzhou IECHO Automatic Technology Co., Ltd Building 11(C1), NO.20 Dongqiao Road, Dongzhou Subdistrict, Fuyang District, Hangzhou city, Zhejiang Province, China. 2) CERTIFICATE NO.: ISETC.001020200528 TECHNICAL REFERENCE: IECHO-2020A01 3) ISET MARK: 4) CAUTION ABOUT CE MARKING (Instruction for the Applicant who puts the product on the EU market): The label of the CE Marking on the left side should be not less than 5mm height. CE Marking and EC Declaration of Conformity are duties for the manufacturer or its applicant who puts the product on the market. This one is responsible to start the CE marking and certification procedure as required by the legislation in force. Only for the products which are compulsorily included into specific Directives or Regulations will be necessary to appoint a Notified Body. 5) TYPE OF PRODUCT: Digital Cutting Machine MODEL(S): BK, BK2, BK3, BKL, BKM, BKMS, GLK, LCP, SC, SCT, TK, TK3S, GLS, LCPS, PK, VK, TK4S, SK2 6) LIST OF DIRECTIVES / REGULATIONS /STANDARDS (as declared by the manufacturer itself) Machinery Directive 2006/42/EC, Low Voltage Directive 2014/35/EU, Electromagnetic Compatibility 2014/30/EU EN 12044:2005+A1:2009; EN 60204-1:2018, EN 61000-6-1:2019, EN 61000-6-3:2007+A1:2011 7) NOTE: This document is not referred to any evaluation that could be considered as included in the scope of the activities covered by the standard BS EN ISO/IEC 17065:2012 or European Regulation 765/2008. -

List of Main Production Facilities of ALDI Nord's Suppliers for Apparel
List of Main Production Facilities of ALDI Nord‘s Suppliers for Apparel, Home Textiles and Shoes Version: April 2021 Produktionsstättenliste | März 2018 | Seite 0/17 Name Address Number of Employees Commodity Group Bangladesh AB Apparels Ltd. 225, Singair Road, Tetuljhora, Hemayetpur 2001 - 5000 Garment textiles Ador Composite Ltd. 1, C & B Bazar, Gilarchala, Sreepur 1001 - 2000 Garment textiles AKH Eco Apparels Ltd. 495, Balitha, Shahbelishwar, Dhamrai 5001 - 10000 Garment textiles Angshuk Ltd. 133-134, Hamayetpur, Savar 501 - 1000 Garment textiles Apparels Village Ltd. Khagan, Birulia, Savar 2001 - 5000 Garment textiles Aspire Garments Ltd. 491, Dhalla, Singair 2001 - 5000 Garment textiles B.H.I.S. Apparels Ltd. 671, Datta Para, Hossain Market, Tongi 2001 - 5000 Garment textiles Blue Planet Knitwear Ltd. Mulaid, P.O.: Tengra, Sreepur 1001 - 2000 Garment textiles Chaity Composite Ltd. Chotto Silmondi, Tripurdi, Sonargaon 5001 - 10000 Garment textiles Chantik Garments Ltd. Kumkumari, Gouripur, Ashulia, Savar 2001 - 5000 Garment textiles Chorka Textile Ltd. Kajirchor, Danga Bazar, Polash 2001 - 5000 Garment textiles Citadel Apparels Ltd. Joy Bangla Road, Kunia, K.B. Bazar, Gazipur Sadar 501 - 1000 Garment textiles Cotton Dyeing & Finishing Mills Ltd. Vill: Amtoli, Union: 10 No. Habirbari, P.O-Seedstore Bazar, P.S.-Valuka 1001 - 2000 Garment textiles Crossline Factory (Pvt) Ltd. 25, Vadam, Uttarpara, Nishatnagar, Tongi 1001 - 2000 Garment textiles Plot No. 45, 48, 49, 51 & 52; Holding No.: 3/C, Vadam, P.O.: Crossline Knit Fabrics Ltd. 1001 - 2000 Garment textiles Nishatnagar, Tongi, Gazipur-1711, Gazipur Crown Exclusive Wears Ltd. Mawna, Sreepur 2001 - 5000 Garment textiles Crown Fashion & Sweater Industries Ltd. Plot No. 781-782, Vogra, Joydebpur, Gazipur-1704 2001 - 5000 Garment textiles Denim Fashions Ltd. -

SUPPLIER LIST AUGUST 2019 Cotton on Group - Supplier List 2
SUPPLIER LIST AUGUST 2019 Cotton On Group - Supplier List _2 COUNTRY FACTORY NAME SUPPLIER ADDRESS STAGE TOTAL % OF % OF % OF TEMP WORKERS FEMALE MIGRANT WORKER WORKERS WORKER CHINA NINGBO FORTUNE INTERNATIONAL TRADE CO LTD RM 805-8078 728 LANE SONGJIANG EAST ROAD SUP YINZHOU NINGBO, ZHEJIANG CHINA NINGBO QIANZHEN CLOTHES CO LTD OUCHI VILLAGE CMT 104 64% 75% 6% GULIN TOWN, HAISHU DISTRICT NINGBO, ZHEJIANG CHINA XIANGSHAN YUFA KNITTING LTD NO.35 ZHENYIN RD, JUEXI STREET CMT 57 60% 88% 12% XIANGSHAN COUNTY NINGBO CITY, ZHEJIANG CHINA SUNROSE INTERNATIONAL CO LTD ROOM 22/2 227 JINMEI BUILDING NO 58 LANE 136 SUP SHUNDE ROAD, HAISHU DISTRICT NINGBO, ZHEJIANG CHINA NINGBO HAISHU WANQIANYAO TEXTILE CO LTD NO 197 SAN SAN ROAD CMT 26 62% 85% 0% WANGCHAN INDUSTRIAL ZONE NINGBO, ZHEJIANG CHINA ZHUJI JUNHANG SOCKS FACTORY DAMO VILLAGE LUXI NEW VILLAGE CMT 73 38% 66% 0% ZHUJI CITY ZHEJIANG CHINA SKYLEAD INDUSTRY CO LIMITED LAIMEI INDUSTRIAL PARK SUP CHENGHAI DISTRICT, SHANTOU CITY GUANGDONG CHINA CHUANGXIANG TOYS LIMITED LAIMEI INDUSTRIAL PARK CMT 49 33% 90% 0% CHENGHAI DISTRICT SHANTOU, GUANGDONG CHINA NINGBO ODESUN STATIONERY & GIFT CO LTD TONGJIA VILLAGE, PANHUO INDUSTRIAL ZONE SUP YINZHOU DISTRICT NINGBO CITY, ZHEJIANG CHINA NINGBO ODESUN STATIONERY & GIFT CO LTD TONGJIA VILLAGE, PANHUO INDUSTRIAL ZONE CMT YINZHOU DISTRICT NINGBO CITY, ZHEJIANG CHINA NINGBO WORTH INTERNATIONAL TRADE CO LTD RM. 1902 BUILDING B, CROWN WORLD TRADE PLAZA SUP NO. 1 LANE 28 BAIZHANG EAST ROAD NINGBO ZHEJIANG CHINA NINGHAI YUEMING METAL PRODUCT CO LTD NO. 5 HONGTA ROAD -

April 2020 ● Investors Newsletter (Stock Code: 00672.HK)
ZHONG AN GROUP 众安集团有限公司 April 2020 ● Investors Newsletter (stock code: 00672.HK) On 21 November 2019,Zhong An Group, through public auction, Land successfully acquired a plot in Longwan district, Wenzhou city, Zhejiang province, at a total consideration of 928 million. The Information plot, with a site area of 43,630 sq.m and a planned GFA of 109,074 sq.m, is for residential and commercial purposes. On 17 January, 2020, Zhong An Group successfully acquired a plot in core area of Wenzhou city, Zhejiang province, at a total consideration of 1.12 billion though public auction. The plot, with a site area of 30,236 sq.m and a planned GFA of 84,964 sq.m,is for residential purpose. On 3 February,2020, Zhong An Group successfully acquired a plot in Fuyang district, Hangzhou city, Zhejiang province, at a total consideration of RMB1.12 billion. The plot, with a site area of 54,493 sq.m and a planned GFA of 130,783 sqm, is for residential purpose. On 24 February, 2020, Zhong An Group successfully acquired a plot in Wenzhou city, Zhejiang province, at a total consideration of 1.14 billion. The plot, with a total site area of 36,230sq.m and a planned GFA of 144,793 sq.m,is for residential purpose. On 5 March,2020,Zhong An Group successfully acquired a plot in Beilun District, Ningbo city, Zhejiang province, at a total consideration of 984 million. The plot, with a site area of 36,230 sq.m and a planned GFA of 79,706 sq.m, is for residential purpose. -

BANK of NINGBO CO., LTD. 2020 Annual Report
BANK OF NINGBO CO., LTD. (Stock Code: 002142) 2020 Annual Report Full Text of 2020 Annual Report of Bank of Ningbo Co., Ltd. Chapter One Important Notes, Content and Interpretation The Board of Directors, Board of Supervisors, directors, supervisors and senior management of the Company ensure the authenticity, accuracy and completeness of contents, and guarantee no frauds, misleading statements or major omissions in this report. They are willing to burden any individual and joint legal responsibilities. The 6th meeting of the 7th Board of Directors of the Company deliberated on and approved the text and abstract of 2020 Annual Report. 13 directors in person out of the total of 13 required directors attended the meeting, and part of supervisors attended as a nonvoting delegates. The Chairman of the Company, Mr. Lu Huayu, the President, Mr. Luo Mengbo, the person in charge of accounting, Mr. Zhuang Lingjun, and the general manager of financial department, Ms. Sun Hongbo hereby declare to guarantee the authenticity, accuracy and completeness of financial statements in the annual report. Financial data and indicators included in this annual report are following the criteria of Chinese Accounting Standard for Business Enterprises. Unless otherwise stated, all data in the consolidated financial statements of Bank of Ningbo Co., Ltd. and its holding subsidiary, Maxwealth Fund Management Co., Ltd., its wholly-owned subsidiaries, Maxwealth Financial Leasing Co., Ltd. and Ningyin Finance Co., Ltd., are subject to the unit of RMB. Ernst & Young Hua Ming LLP audited the 2020 Financial Statements of the Company in accordance with domestic accounting principles and published unqualified opinion. -

中國交通建設股份有限公司 China Communications
Hong Kong Exchanges and Clearing Limited and The Stock Exchange of Hong Kong Limited take no responsibility for the contents of this announcement, make no representation as to its accuracy or completeness and expressly disclaim any liability whatsoever for any loss howsoever arising from or in reliance upon the whole or any part of the contents of this announcement. 中國交通建設股份有限公司 CHINA COMMUNICATIONS CONSTRUCTION COMPANY LIMITED (A joint stock limited company incorporated in the People’s Republic of China with limited liability) (Stock Code: 1800) ANNOUNCEMENT OF ANNUAL RESULTS FOR THE YEAR ENDED 31 DECEMBER 2020 FINANCIAL HIGHLIGHTSNote Revenue of the Group in 2020 amounted to RMB624,495 million, representing an increase of RMB71,381 million or 12.9% from RMB553,114 million in 2019. Gross profit in 2020 amounted to RMB80,036 million, representing an increase of RMB10,739 million or 15.5% from RMB69,297 million in 2019. Operating profit in 2020 amounted to RMB34,405 million, representing an increase of RMB273 million or 0.8% from RMB34,132 million in 2019. Profit before tax in 2020 amounted to RMB26,957 million, compared with RMB27,349 million in 2019. Profit attributable to owners of the parent in 2020 amounted to RMB16,475 million, compared with RMB19,999 million in 2019. Earnings per share for the year 2020 amounted to RMB0.92, compared with RMB1.16 for the year 2019. The value of new contracts of the Group in 2020 amounted to RMB1,066,799 million, representing an increase of 10.6% from RMB964,683 million in 2019. As at 31 December 2020, the backlog for the Group was RMB2,910,322 million. -

FACTORY LIST Tier 1 Trade Partners
FACTORY LIST Tier 1 Trade Partners Product Parent Company Facility Name Facility Address Province Country Number of Category Workers Apparel Calzificio Telemaco SRL Calzificio Telemaco SRL Via Brentella 9, Cap 31040, Trevignano Treviso Italy < 50 Accessories B.S.T. International Trade Company Ltd Chanchang Stainless Steel Products Co., Ltd Xiangzhu Industrial Zone, Yongkang City 322300 Zhejiang China < 50 Apparel Changsu Gloves Factory Co. Changsu Gloves Factory Co. No. 199, Xichun Lane, Hupu Lane, Bixi New District, Changshu City 2115500 Jiangsu China < 50 Apparel Changsu Haozu Socks Co., Ltd Changsu Haozu Socks Co., Ltd No. 6, Minda Road, XushiQigan Village, Dongbin Town, Changshu City 2115500 Jiangsu China < 50 Apparel Castlecrafts Ltd T/A Dewhirst Menswear Dalian Taiyangying Garments Co.,Ltd Chengguan Community, Pikou Town Sub-district, Pulandian District, Dalian Liaoning China 200 - 300 Accessories Delta (Tien Giang) Co., Ltd Delta (Tien Giang) Co., Ltd 69B&70A, Long jiang Industrial Park, Tan Lap 1 valley, Tan Phuoc County, Tiền Giang Vietnam 100 - 200 Accessories Favour Light Co.,Ltd DongGuan Sheng Shi Hardware & Plastics Prod. Co. Ltd No.3 Daling Street, Jiao Yi Tang Administrative District, Tang Xia Town Guangdong China 800 - 900 Accessories Dongguan Yifeng Lock Co. Ltd Dongguan Yifeng Lock Co. Ltd Building A-H, No.28, Street 6, First Gongye Road, Tutang Village, Changping Town, Dongguan Guangdong China 1100 - 1200 Apparel DongYang New DongHuang Garment Co., Ltd DongYang New DongHuang Garment Co., Ltd No.52 Jinwang Road, West -

Como Parte Fundamental Del Compromiso De MANGO Con La
LISTA DE FÁBRICAS DE PRODUCCIÓN DE MANGO Como parte fundamental del compromiso de MANGO con la sostenibilidad MANGO da respuesta así al compromiso adquirido con CCOO Industria en es esencial establecer una gestión responsable de nuestra cadena de el “Acuerdo Bilateral Global para garantizar las normas internacionales en la suministro que, junto con la participación de determinados grupos de interés, cadena de suministro de MANGO” y a los requerimientos del Transparency contribuye a la transparencia y es clave en la debida diligencia hacia Pledge Standard, que insta a las marcas a hacer público el nombre, dirección nuestros proveedores. Priorizamos una relación de confianza con ellos, y otros detalles de las fábricas de producción. asegurando que están alineados con nuestros propios compromisos, Cabe destacar que MANGO no posee ninguna fábrica en propiedad y contribuyendo así a asegurar los derechos humanos de los trabajadores y a ninguna trabaja en exclusiva para nuestra organización. Algunas de ellas fortalecer la industria textil a nivel global. pueden utilizarse de forma discontinua en el tiempo en función de las Así pues, conscientes de la importancia de ser transparentes y de facilitar el características del producto por lo que la lista se actualizará mínimo dos acceso a la información para asegurar este compromiso, hacemos pública a veces al año. Asimismo, se irán ampliando progresivamente los datos través de nuestra página web la lista de fábricas de nivel 1 (tier 1) que han aportados sobre nuestra cadena de suministro. producido para MANGO en el presente año 2020. País Fábrica Dirección Ciudad Categoría Nº Empleados ALBANIA DONA ELEGANT SHPK Rruga Vangjel Noti, Ambiente Edil Tirane Complementos < 1000 Centro, Kapanoni 1, Uz 2000, 1029 BANGLADESH A.K.M.