Population Structure of African Mahoganies in Four Forest Reserves: Implications for Conservation and Management in Ghana
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

African Mahogany Anegre Birdseye Maple Black Walnut
African Mahogany African Mahogany (Khaya) is a genus of seven species of trees in the mahogany family Meliaceae, native to tropical Africa and Madagascar. All species become big trees 30-35 m tall, rarely 45 m, with a trunk over 1 m trunk diameter, often buttressed at the base. The leaves are pinnate, with 4-6 pairs of leaflets, the terminal leaflet absent; each leaflet is 10-15 cm long abruptly rounded toward the apex but often with an acuminate tip. The leaves can be either deciduous or evergreen depending on the species. The flowers are produced in loose inflorescences, each flower small, with four or five yellowish petals and ten stamens. The timber of Khaya is called African mahogany, the only timber widely accepted as mahogany besides that of the true mahogany, of the genus Swietenia. Khaya senegalensis, also known as the African dry zone mahogany is also used for its non timber parts. In West Africa, Fulani herdsmen prune the tree during the dry season to feed cattle. Anegre Anegre is milled from the Tawa tree (Beilschmiedia tawa) is a New Zealand broadleaf tree common in the central parts of the country. Tawa is often the dominant canopy species in lowland forests in the North Island and north east of the South Island, Individual specimens may grow up to 30 meters or more in height with trunks up to 1.2 meters in diameter, and they have smooth dark bark. The word "tawa" is the Maori name for the tree. One of the few hardwood trees in the country with good timber, the wood of this tree can be used for attractive and resilient floor boarding. -

COMPARATIVE ECOLOGICAL WOOD ANATOMY of AFRICAN MAHOGANY KHAYA IVORENSIS with SPECIAL REFERENCE to DAMAGE CAUSED by HYPSIPYL ROBUSTA SHOOTBORER Rinne E., Hakkarainen J
Структурные и функциональные отклонения от нормального роста и развития растений COMPARATIVE ECOLOGICAL WOOD ANATOMY OF AFRICAN MAHOGANY KHAYA IVORENSIS WITH SPECIAL REFERENCE TO DAMAGE CAUSED BY HYPSIPYL ROBUSTA SHOOTBORER Rinne E., Hakkarainen J. A., Rikkinen J. Department of Biosciences, University of Helsinki, PO Box 65, FI-00014 University of Helsinki, Finland, E-mail: [email protected] Abstract. This study focuses on the wood anatomy of 60 Khaya ivorensis A. Chev. (Meliaceae) trees from natural forests in three forest zones of southern Ghana. The arrangement, grouping, density and diameter of vessel elements in stem wood were measured and theoretical hydraulic conductivities were calculated for each site. Also anatomical characteristics of thin branches damaged by mahogany shootborer (Hypsipyla robusta Moore) were studied with special reference to effects on vessel element diameter, vessel density and hydraulic conductivity. Our study revealed that there were significant differences in vessel element diameters and hydraulic conductivities between the sites, and that the water conduction efficiency of K. ivorensis wood increased with increasing annual precipitation. In shootborer damaged branches mean vessel element diameters were consistently smaller and mean theoretical hydraulic conductivities lower than in undamaged branches. This suggests that Hypsipyla damage can have a negative effect on the growth rates of mature African mahogany trees in natural forests. Introduction. Khaya ivorensis A. Chev. is distributed throughout the moist lowlands of tropical West Africa. The species has a wide habitat tolerance, but it favors the banks of rivers and streams. The wood is highly valued especially for furniture, cabinet work and light flooring [10]. The species has become commercially extinct in large parts of its original range. -

African Mahogany Wood Defects Detected by Ultrasound Waves
General Technical Report FPL-GTR-239 • Proceedings: 19th International Nondestructive Testing and Evaluation of Wood Symposium African mahogany wood defects detected by ultrasound waves Tamara Suely Filgueira Amorim França Department of Sustainable Bioproducts, Mississippi State University, Starkville, Mississippi, United States, [email protected] Frederico Jose Nistal França Department of Sustainable Bioproducts, Mississippi State University, Starkville, Mississippi, United States, [email protected] Robert John Ross U.S. Forest Service, Forest Product Laboratory, Madison, Wisconsin, USA, [email protected] Xiping Wang U.S. Forest Service, Forest Product Laboratory, Madison, Wisconsin, USA, [email protected] Marina Donaria Chaves Arantes Departamento de Ciências Florestais e da Madeira, Universidade Federal do Espírito Santo, Jerônimo Monteira, Espírito Santo, Brasil, [email protected] Roy Daniel Seale Department of Sustainable Bioproducts, Mississippi State University, Starkville, Mississippi, United States, [email protected] Abstract This study aims to investigate the potential of ultrasound wave to detect defects in 19 years old of two species of African mahogany planted in Brazil. Were used five 76 x 5 x 5 cm samples from each species with different types of defects, and were conditioned to 12% moisture content. The samples were scanned with ultrasound wave in longitudinal direction and every 2,54 cm in radial and tangential directions along the samples. It was possible to identify end split and pin knots in Khaya ivorensis and reaction wood in Khaya senegalensis wood. Beetle galleries did not affect wave velocities in Khaya senegalensis wood. Grain angle had a large effect in ultrasound velocities in radial and tangential directions. Khaya senegalensis exhibit lower longitudinal velocities related to larger amount of interlocked grain in this species. -

Lumber / Veneer Match
Lumber Veneer Type of Cut Alder (Select) Alder Plain Sliced Face Alder (Knotty) Knotty Alder (Dark Putty in open knots) Plain Sliced Face Ash (Eastern White) Ash (Eastern White) Rotary Multi Piece Face Ash (Western) Ash (Eastern White) Rotary Multi Piece Face Beech (European Steamed) Beech (European Steamed) Plain Sliced Face Birch (Eastern Red) N/A N/A Birch (Eastern White) Birch (Eastern White) Rotary Multi Piece Face Birch (Western) N/A Cherry (Select) Cherry (Select) Plain Sliced Face Cherry (Rustic) Fir (CVG) Fir (CVG) Plain Sliced Face Hickory (Brown Heart) Hickory (Brown Heart) Plain Sliced Face Hickory (Calico) Hickory (Calico) Plain Sliced Face Hickory (Rustic Brown Heart) Hickory (Rustic Brown Heart) Plain Sliced Face Hickory (Rustic Calico) Hickory (Rustic Calico) Plain Sliced Face Lyptus Lyptus Plain Sliced Face Mahogany (African Khaya) Mahogany (African Khaya) Plain Sliced Face Mahogany (Edinam) Mahogany (African Khaya) Plain Sliced Face Maple (Eastern Hard White) Maple (Eastern Hard White) Rotary Whole Piece Face Maple (Eastern Soft Rustic) Maple (Eastern Hard Rustic) Plain Sliced Face Maple (Eastern Soft White) Maple (Eastern Hard White) Rotary Whole Piece Face Maple (Western) Alder Plain Sliced Face Paint Grade (Lumber Panel) DCD's Choice Rotary Whole Piece Face Paint Grade (MDF Panel) MDF N/A Pine (Eastern Clear) Pine (Eastern Clear) Plain Sliced Face Pine (Eastern Knotty) Pine (Eastern Knotty) Plain Sliced Face Poplar N/A N/A Red Oak (Natural) Red Oak (Natural) Plain Sliced Face Red Oak (Qtr Sawn) Red Oak (Qtr Sawn) Quarter Sawn Face Red Oak (Rift) Red Oak (Rift) Rift Sawn Face Red Oak (Select) Red Oak (Select) Plain Sliced Face Sapele (Flat Sawn) Sapele (Flat Sawn) Plain Sliced Face Sapele (Qtr Sawn) Sapele (Qtr Sawn) Quarter Sawn Face Walnut Walnut Plain Sliced Face White Oak (Select) White Oak (Select) Plain Sliced Face White Oak (Qtr Sawn) White Oak (Qtr Sawn) Quarter Sawn Face White Oak (Rift) White Oak (Rift) Rift Sawn Face. -

Luthier's Dictionary
LUTHIER’S DICTIONARY ©2014 The European Guitar Builders Association ©2014 The European Guitar Builders Association Woods ENGLISH DUTCH GERMAN FINNISH FRENCH ITALIAN SPANISH SWEDISH LATIN Alder Elzen Erle Leppä Aulne Ontano Aliso Al Alnus Alder / european, black, Elzen / Europees, Erle Leppä / eurooppalainen Aulne européen Ontano europeo Aliso Europeo Al (Klibbal) Alnus glutinosa common zwart, gewoon Alder / red, western red Elzen Erle Leppä / amerikkalainen Aulne rouge, Aulne Ontano americano Aliso Rojo Rödal Alnus rubra américain Arctic Birch Berken Birke Jääkoivu Bouleau arctique Betulla artica (bianca) Abedul del Ártico Björk Betula pendula Ash Essen Esche Saarni Frêne Frassino Fresno Ask Fraxinus Ash / american Amerikaans essen Esche Saarni / amerikkalainen Frêne américain Frassino americano Fresno Americano Vitask Fraxinus americana Ash / European Europees essen Esche Saarni / eurooppalainen Frêne européen Frassino europeo Fesno europeo Europeisk ask Fraxinus excelsior Ash / Swamp Ash Moeras essen Sumpfesche Suosaarni Frêne des marais Frassino palustre Fesno del Pantano Svartask Fraxinus nigra Basswood Linden Linde Lehmus Tilleul Tiglio Tilo Lind Tilia americana Bubinga Bubinga Bubinga Bubinga Bubinga Bubinga Bubinga Bubinga Guibourtia spp. (G. demeusei, G. pellegriniana, G. tessmannii, etc.) Cocobolo Cocobolo Cocobolo Cocobolo Coccobolo Cocobolo Cocobolo Mexikansk jakaranda Dalbergia retusa Ebony / African Afrikaans ebben Ebenholz Eebenpuu / Ébène Ebano africano Ébano Africano Ebenholts Diospyros crassiflora afrikkalainen Ebony / -
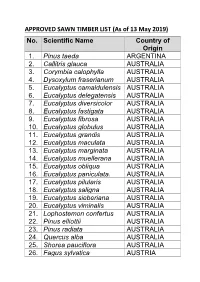
APPROVED SAWN TIMBER LIST (As of 13 May 2019) No. Scientific Name Country of Origin 1
APPROVED SAWN TIMBER LIST (As of 13 May 2019) No. Scientific Name Country of Origin 1. Pinus taeda ARGENTINA 2. Callitris glauca AUSTRALIA 3. Corymbia calophylla AUSTRALIA 4. Dysoxylum fraserianum AUSTRALIA 5. Eucalyptus camaldulensis AUSTRALIA 6. Eucalyptus delegatensis AUSTRALIA 7. Eucalyptus diversicolor AUSTRALIA 8. Eucalyptus fastigata AUSTRALIA 9. Eucalyptus fibrosa AUSTRALIA 10. Eucalyptus globulus AUSTRALIA 11. Eucalyptus grandis AUSTRALIA 12. Eucalyptus maculata AUSTRALIA 13. Eucalyptus marginata AUSTRALIA 14. Eucalyptus muellerana AUSTRALIA 15. Eucalyptus obliqua AUSTRALIA 16. Eucalyptus paniculata. AUSTRALIA 17. Eucalyptus pilularis AUSTRALIA 18. Eucalyptus saligna AUSTRALIA 19. Eucalyptus sieberiana AUSTRALIA 20. Eucalyptus viminalis AUSTRALIA 21. Lophostemon confertus AUSTRALIA 22. Pinus elliottii AUSTRALIA 23. Pinus radiata AUSTRALIA 24. Quercus alba AUSTRALIA 25. Shorea pauciflora AUSTRALIA 26. Fagus sylvatica AUSTRIA No. Scientific Name Country of Origin 27. Picea abies AUSTRIA 28. Picea abies BELARUS 29. Pinus sylvestris BELARUS 30. Quercus alba BELGIUM 31. Dipteryx odorata BOLIVIA 32. Apuleia leiocarpa BRAZIL 33. Astronium lecointei BRAZIL 34. Bagassa guianensis BRAZIL 35. Cedrela odorata BRAZIL 36. Cedrelinga catenaeformis BRAZIL 37. Couratari guianensis BRAZIL 38. Dipteryx odorata BRAZIL 39. Eucalyptus grandis BRAZIL 40. Eucalyptus grandis BRAZIL 41. Hymenaea courbaril BRAZIL 42. Hymenolobium modestum BRAZIL 43. Hymenolobium Nitidum BRAZIL Benth 44. Hymenolobium BRAZIL pulcherrimum 45. Manilkara bidentata BRAZIL 46. Myroxylon balsamum BRAZIL 47. Pinus radiata BRAZIL 48. Pinus taeda BRAZIL 49. Quassia simarouba BRAZIL 50. Tectona grandis BRAZIL 51. Fagus sylvatica BULGARIA No. Scientific Name Country of Origin 52. Quercus alba BULGARIA 53. Chlorophora excelsa CAMEROON 54. Cylicodiscus gabunensis CAMEROON 55. Entandrophragma CAMEROON candollei 56. Entandrophragma CAMEROON cylindricum 57. Entandrophragma CAMEROON cylindricum 58. Entandrophragma utile CAMEROON 59. -

Federal Trade Commission § 243.4
Federal Trade Commission § 243.4 (2) Under this section, unqualified usage of that term. Examples of im- phrases such as ``walnut'', ``walnut fin- proper use of the term ``mahogany'' in- ish'', ``in walnut'', ``fruitwood'', ``oak'', clude reference to Red Lauan as ``mahogany finish'', and other terms of ``Lauan mahogany'' or to White Lauan similar import or meaning, will not be as ``Blond Lauan mahogany''. Such adequate. But statements such as woods, however, may be described as ``walnut stain'', ``maple stain finish'', ``Red Lauan'' or ``Lauan'' or ``White ``mahogany finish on gum'', ``photo- Lauan'', respectively. The term ``Phil- graphically reproduced pecan grain'', ippine mahogany'' will be accepted as a ``printed pecan design'', ``fruitwood fin- name or designation of the seven woods ish on selected hardwood veneer'', named above. Such term shall not be ``cherry grain finish on vinyl overlay'' applied to any other wood, whether or and ``walnut finish on other hard- not grown on the Philippine Islands. woods'' (or ``softwoods'', as the case (4) The term ``mahogany'', with or may be) will satisfy this provision if without qualifications, should not be such statements are factually correct and appear in contexts which are oth- used to describe any other wood except erwise nondeceptive. 2 [Guide 2] as provided above. This applies also to any of the woods belonging to the § 243.3 Deceptive use of wood names. Meliaceae family, other than genera Industry members should not use any Swietenia and Khaya. direct or indirect representation con- (c) Maple. The terms hard maple, rock cerning the identity of the wood in in- maple, bird's-eye maple, Northern maple dustry products that is false or likely or other terms of similar nature should to mislead purchasers as to the actual not be used to describe woods other wood composition. -

Chapter 3--Physical Properties and Moisture Relations of Wood
Chapter 3 Physical Properties and Moisture Relations of Wood William Simpson and Anton TenWolde he versatility of wood is demonstrated by a wide Contents variety of products. This variety is a result of a Appearance 3–1 spectrum of desirable physical characteristics or properties among the many species of wood. In many cases, Grain and Texture 3–1 more than one property of wood is important to the end Plainsawn and Quartersawn 3–2 product. For example, to select a wood species for a product, the value of appearance-type properties, such as texture, grain Decorative Features 3–2 pattern, or color, may be evaluated against the influence of Moisture Content 3–5 characteristics such as machinability, dimensional stability, Green Wood and Fiber Saturation Point 3–5 or decay resistance. Equilibrium Moisture Content 3–5 Wood exchanges moisture with air; the amount and direction of the exchange (gain or loss) depend on the relative humid- Sorption Hysteresis 3–7 ity and temperature of the air and the current amount of water Shrinkage 3–7 in the wood. This moisture relationship has an important Transverse and Volumetric 3–7 influence on wood properties and performance. This chapter discusses the physical properties of most interest in the Longitudinal 3–8 design of wood products. Moisture–Shrinkage Relationship 3–8 Some physical properties discussed and tabulated are influ- Weight, Density, and Specific Gravity 3–11 enced by species as well as variables like moisture content; Working Qualities 3–15 other properties tend to be independent of species. The thor- oughness of sampling and the degree of variability influence Decay Resistance 3–15 the confidence with which species-dependent properties are Thermal Properties 3–15 known. -

Sequestered Standing Carbon Stock in Selective Exploited Timbers
e-ISSN:2320-0189 p-ISSN:2347-2308 Research & Reviews: Journal of Botanical Sciences Sequestered Standing Carbon Stock in Selective Exploited Timbers Grown in Tropical Forest: a Case Study from the National Park of Lobeke (Cameroon) Noiha Noumi Valery1*, Zapfack Louis2, Ngueguim Jules Romain3, Tabue Mbobda Roger Bruno2, Ibrahima Adamou1 and Mapongmetsem Pierre Marie1 1University of Ngaoundere, Faculty of Science, Department of Biological Sciences, Laboratory of Biodiversity and Sustainable Development. P.O. Box. 454 Ngaoundere, Cameroon. 2University of Yaounde I, Faculty of Science, Department of Plant Biology, Laboratory of Plant Systematic and Ecology, P.O. Box. 812 Yaounde, Cameroon. 3Institute of Agricultural Research for Development (IRAD), P.O. Box 219 Kribi, Cameroon. Research Article Received date: 26/06/2015 ABSTRACT Accepted date: 10/09/2015 The study was carried out in the National Park of Lobeke (Cameroon). Published date: 12/09/2015 It aimed to assess diversity and sequestration potential of some commercialized timbers under industrialisation pressure. We took place *For Correspondence 37 transects of 5 ha each. All selected timbers of at least 5 cm of DBH were surveyed. 8, 619 individuals divided into 6 families, 6 genera and Noiha Noumi Valery, University of Ngaoundere, 9 species were recorded. Diospyros crassiflora (Ebene) was the most Faculty of Science, Department of Biological diversified species (ISH= 8.09); followed Terminalia superba (Frake; ISH= Sciences, Laboratory of Biodiversity and 2.08). Triplochyton scleroxylon (Ayous), Terminalia superba (Frake) and Sustainable Development. P.O. Box. 454 Entandrophragma cylindricum (Sapelli) were the biggest species of the Ngaoundere, Cameroon. Tel: +237 670 835 813 flora with 549.01; 370.22 and 334.8 m² of basal area respectively. -

Harmonized Tariff Schedule of the United States (2020) Revision 26 Annotated for Statistical Reporting Purposes
Harmonized Tariff Schedule of the United States (2020) Revision 26 Annotated for Statistical Reporting Purposes SECTION IX WOOD AND ARTICLES OF WOOD; WOOD CHARCOAL; CORK AND ARTICLES OF CORK; MANUFACTURES OF STRAW, OF ESPARTO OR OF OTHER PLAITING MATERIALS; BASKETWARE AND WICKERWORK IX-1 Harmonized Tariff Schedule of the United States (2020) Revision 26 Annotated for Statistical Reporting Purposes IX-2 Harmonized Tariff Schedule of the United States (2020) Revision 26 Annotated for Statistical Reporting Purposes CHAPTER 44 WOOD AND ARTICLES OF WOOD; WOOD CHARCOAL IX 44-1 Notes 1. This chapter does not cover: (a) Wood, in chips, in shavings, crushed, ground or powdered, of a kind used primarily in perfumery, in pharmacy, or for insecticidal, fungicidal or similar purposes (heading 1211); (b) Bamboo or other materials of a woody nature of a kind used primarily for plaiting, in the rough, whether or not split, sawn lengthwise or cut to length (heading 1401); (c) Wood, in chips, in shavings, ground or powdered, of a kind used primarily in dyeing or in tanning (heading 1404); (d) Activated charcoal (heading 3802); (e) Articles of heading 4202; (f) Goods of chapter 46; (g) Footwear or parts thereof of chapter 64; (h) Goods of chapter 66 (for example, umbrellas and walking-sticks and parts thereof); (ij) Goods of heading 6808; (k) Imitation jewelry of heading 7117; (l) Goods of section XVI or section XVII (for example, machine parts, cases, covers, cabinets for machines and apparatus and wheelwrights© wares); (m) Goods of section XVIII (for -

The Difference
Products that deliver the difference UTAH DIVISION Decorative Surfaces TFL StyleLite High Gloss and Matte Panels are the superior choice for creating stunning, modern interior spaces. StyleLite delivers a superior appearance to high pressure laminate, TFL, paint, PVC and other thermfoil faced panels. Super Matte panels are manufactured by the application of matte foil to It offers cabinetmakers, millworkers, furniture makers and the interior the refined and calibrated MDF. Smooth and velvety surface design design industry a high gloss solution that is both luxurious and affordable. creates a warm, cozy ambience. Durable. Stylish. Affordable. Timber Products’ TFL is available in a wide Ultra High Gloss (UHG) is made by laminating a very durable decorative range of colors, woodgrains, and decorative patterns. Using your choice of foil on a refined calibrated and one-sided TFL coated MDF. Without the high-quality substrates, most manufactured in their own facilities. They can need to take protective measures, UHG boards are ready for use and easy also custom-design and engineer a panel to meet your exact specifications. to process: simply cut, edge and use. Offering a luxurious collection of woodgrains, exotics and linear designs VelourTouchTM captures today’s trends of extreme matte finishes and that are versatile enough to transcend market segments and geographic painted cabinet looks. Quality, beautiful, worry-free and affordable regions. The strength of Uniboard® TFL goes beyond quality to capture materials are manufactured by Olon just for your building needs. The the perfect balance between commercial and residential design. VelourTouchTM surface is silky to the touch, anti-fingerprint and scratch resistant. -

Inventory of Natural Regeneration and the Recovery of Logging Gaps In
UNIVERSITY OF APPLIED SCIENCES VAN HALL LARENSTEIN Inventory of Natural Regeneration and the Recovery of Logging Gaps in the Nkrabia Forest Reserve in Ghana A Comparison between Chainsaw Milling and Conventional Logging Thorsten Dominik Herrmann Bachelor Thesis to obtain the academic degree Bachelor of Science in Forestry and Bachelor in Forest and Nature Management at the University of Applied Sciences Van Hall Larenstein, Part of Wageningen University and Research Center Topic: Inventory of Natural Regeneration and the Recovery of Logging Gaps in the Nkrabia Forest Reserve in Ghana – A Comparison between Chainsaw Milling and Conventional Logging Author: Thorsten Dominik Herrmann Rieslingstraße 29 71364 Winnenden, Germany [email protected] 1. Examinor: Jaap de Vletter 2. Examinor: Mr. Zambon External supervisor: Robbert Wijers Closing date: Velp, 03 / January 2011 Key words: Ghana, natural regeneration, logging gaps, chainsaw milling ACKNOWLEDGEMENT This study was carried out on the behalf of Houthandel Wijers BV and the Forestry Research Institute of Ghana (FORIG) between March and November 2010. The presented research is a continuation of several forestry related studies that had been carried out in the Nkrabia forest reserve in Ghana by students of the University of Applied Sciences Van Hall Larenstein. My special thanks goes to the Forestry Research Institute of Ghana (FORIG) and Robbert Wijers (Houthandel Wijers BV) for giving me the opportunity to conduct the field work in Ghana by financial means and technical as well as logistical support. Furthermore I would like to thank Jonathan Dabo (Technician, FORIG) and Seth Kankam Nuamah (Service Personnel, FORIG) for their indispensable and sedulous support during the field work and data collection for this study.