Fibroblast Growth Factor 2 Regulates Endothelial Cell Sensitivity to Sunitinib
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

ARTICLES Fibroblast Growth Factors 1, 2, 17, and 19 Are The
0031-3998/07/6103-0267 PEDIATRIC RESEARCH Vol. 61, No. 3, 2007 Copyright © 2007 International Pediatric Research Foundation, Inc. Printed in U.S.A. ARTICLES Fibroblast Growth Factors 1, 2, 17, and 19 Are the Predominant FGF Ligands Expressed in Human Fetal Growth Plate Cartilage PAVEL KREJCI, DEBORAH KRAKOW, PERTCHOUI B. MEKIKIAN, AND WILLIAM R. WILCOX Medical Genetics Institute [P.K., D.K., P.B.M., W.R.W.], Cedars-Sinai Medical Center, Los Angeles, California 90048; Department of Obstetrics and Gynecology [D.K.] and Department of Pediatrics [W.R.W.], UCLA School of Medicine, Los Angeles, California 90095 ABSTRACT: Fibroblast growth factors (FGF) regulate bone growth, (G380R) or TD (K650E) mutations (4–6). When expressed at but their expression in human cartilage is unclear. Here, we deter- physiologic levels, FGFR3-G380R required, like its wild-type mined the expression of entire FGF family in human fetal growth counterpart, ligand for activation (7). Similarly, in vitro cul- plate cartilage. Using reverse transcriptase PCR, the transcripts for tivated human TD chondrocytes as well as chondrocytes FGF1, 2, 5, 8–14, 16–19, and 21 were found. However, only FGF1, isolated from Fgfr3-K644M mice had an identical time course 2, 17, and 19 were detectable at the protein level. By immunohisto- of Fgfr3 activation compared with wild-type chondrocytes and chemistry, FGF17 and 19 were uniformly expressed within the showed no receptor activation in the absence of ligand (8,9). growth plate. In contrast, FGF1 was found only in proliferating and hypertrophic chondrocytes whereas FGF2 localized predominantly to Despite the importance of the FGF ligand for activation of the resting and proliferating cartilage. -

Antitumor Reactivity in Vitro Sorafenib Differentially Affect NK Cell The
The Kinase Inhibitors Sunitinib and Sorafenib Differentially Affect NK Cell Antitumor Reactivity In Vitro This information is current as Matthias Krusch, Julia Salih, Manuela Schlicke, Tina of September 24, 2021. Baessler, Kerstin Maria Kampa, Frank Mayer and Helmut Rainer Salih J Immunol 2009; 183:8286-8294; ; doi: 10.4049/jimmunol.0902404 http://www.jimmunol.org/content/183/12/8286 Downloaded from References This article cites 44 articles, 21 of which you can access for free at: http://www.jimmunol.org/content/183/12/8286.full#ref-list-1 http://www.jimmunol.org/ Why The JI? Submit online. • Rapid Reviews! 30 days* from submission to initial decision • No Triage! Every submission reviewed by practicing scientists • Fast Publication! 4 weeks from acceptance to publication by guest on September 24, 2021 *average Subscription Information about subscribing to The Journal of Immunology is online at: http://jimmunol.org/subscription Permissions Submit copyright permission requests at: http://www.aai.org/About/Publications/JI/copyright.html Email Alerts Receive free email-alerts when new articles cite this article. Sign up at: http://jimmunol.org/alerts The Journal of Immunology is published twice each month by The American Association of Immunologists, Inc., 1451 Rockville Pike, Suite 650, Rockville, MD 20852 Copyright © 2009 by The American Association of Immunologists, Inc. All rights reserved. Print ISSN: 0022-1767 Online ISSN: 1550-6606. The Journal of Immunology The Kinase Inhibitors Sunitinib and Sorafenib Differentially Affect NK Cell Antitumor Reactivity In Vitro1 Matthias Krusch,2 Julia Salih,2 Manuela Schlicke,2 Tina Baessler, Kerstin Maria Kampa, Frank Mayer,3 and Helmut Rainer Salih3,4 Sunitinib and Sorafenib are protein kinase inhibitors (PKI) approved for treatment of patients with advanced renal cell cancer (RCC). -

And Insulin-Like Growth Factor-I (IGF-I) in Regulating Human Erythropoiesis
Leukemia (1998) 12, 371–381 1998 Stockton Press All rights reserved 0887-6924/98 $12.00 The role of insulin (INS) and insulin-like growth factor-I (IGF-I) in regulating human erythropoiesis. Studies in vitro under serum-free conditions – comparison to other cytokines and growth factors J Ratajczak, Q Zhang, E Pertusini, BS Wojczyk, MA Wasik and MZ Ratajczak Department of Pathology and Laboratory Medicine, University of Pennsylvania School of Medicine, Philadelphia, PA, USA The role of insulin (INS), and insulin-like growth factor-I (IGF- has been difficult to assess. The fact that EpO alone fails to I) in the regulation of human erythropoiesis is not completely stimulate BFU-E in serum-free conditions, but does do in understood. To address this issue we employed several comp- lementary strategies including: serum free cloning of CD34؉ serum containing cultures indicates that serum contains some cells, RT-PCR, FACS analysis, and mRNA perturbation with oli- crucial growth factors necessary for the BFU-E development. godeoxynucleotides (ODN). In a serum-free culture model, both In previous studies from our laboratory, we examined the ؉ INS and IGF-I enhanced survival of CD34 cells, but neither of role of IGF-I12 and KL9,11,13 in the regulation of early human these growth factors stimulated their proliferation. The influ- erythropoiesis. Both of these growth factors are considered to ence of INS and IGF-I on erythroid colony development was be crucial for the BFU-E growth.3,6,8,14 Unexpectedly, that dependent on a combination of growth factors used for stimul- + ating BFU-E growth. -

(12) Patent Application Publication (10) Pub. No.: US 2009/0111743 A1 Takahashi (43) Pub
US 200901 11743A1 (19) United States (12) Patent Application Publication (10) Pub. No.: US 2009/0111743 A1 Takahashi (43) Pub. Date: Apr. 30, 2009 (54) CYSTEINE-BRANCHED HEPARIN-BINDING Related U.S. Application Data GROWTH FACTOR ANALOGS (60) Provisional application No. 60/656,713, filed on Feb. 25, 2005. (75) Inventor: Kazuyuki Takahashi, O O Germantown, MD (US) Publication Classification (51) Int. Cl. Correspondence Address: A638/08C07K 700 30.82006.O1 201 THIRD STREET, N.W., SUITE 1340 A638/16 (2006.01) ALBUQUEROUE, NM 87102 (US) A638/10 (2006.01) A638/00 (2006.01) (73) Assignee: BioSurface Engineering (52) U.S. Cl. ........... 514/12:530/330; 530/329; 530/328; Technologies, Inc., Rockville, MD 530/327: 530/326; 530/325; 530/324; 514/17; (US) 514/16; 514/15: 514/14: 514/13 (57) ABSTRACT (21) Appl. No.: 11/362,137 The present invention provides a heparin binding growth factor analog of any of formula I-VIII and methods and uses (22) Filed: Feb. 23, 2006 thereof. US 2009/011 1743 A1 Apr. 30, 2009 CYSTENE-BRANCHED HEPARN-BINDING 0007 HBGFs useful in prevention or therapy of a wide GROWTH FACTOR ANALOGS range of diseases and disorders may be purified from natural sources or produced by recombinant DNA methods, however, CROSS-REFERENCE TO RELATED Such preparations are expensive and generally difficult to APPLICATIONS prepare. 0008. Some efforts have been made to generate heparin 0001. This application claims priority to and the benefit of binding growth factor analogs. For example, natural PDGF the filing of U.S. Provisional Patent Application Ser. -

Imatinib-Induced Interstitial Lung Disease and Sunitinib-Associated
CASE Imatinib-induced interstitial lung disease and REPORT sunitinib-associated intra-tumour haemorrhage Herbert H Loong 龍浩鋒 Winnie Yeo 楊明明 An ethnically Chinese patient with newly diagnosed metastatic gastro-intestinal stromal tumour initially treated with imatinib mesylate developed severe interstitial lung disease. As his condition improved after cessation of imatinib mesylate and treatment with corticosteroids, he was started on sunitinib malate. His clinical course was then unfortunately complicated with intra-tumour bleeding. This case report illustrates the dilemmas and complexities associated with treating patients with gastro-intestinal stromal tumours with the new tyrosine kinase inhibitors. Case report A 63-year-old man was referred to our department in January 2007 after being diagnosed with a recurrent gastro-intestinal stromal tumour (GIST). He was initially diagnosed with a duodenal GIST in January 2000 on presenting with symptoms of anaemia. A workup, including upper endoscopy, revealed an ulcerative growth over the third and fourth part of the duodenum. A computed tomographic (CT) scan showed a 4.5 x 5.5 cm soft tissue mass over the same area. A duodenectomy and duodeno-jejunostomy were performed and a pathological examination of tissue removed at surgery confirmed a low-grade GIST (S-100 positive; 4.5 cm in size, mitosis 6/10 high-power field, c-KIT positive). The resection margins were clear so he was managed with routine follow-up and observation. An abdominal ultrasound performed in 2003 showed no evidence of metastases. He remained well until January 2007 when hepatomegaly was found during a physical examination. An abdominal CT scan showed multiple hypervascular tumour foci with cystic changes in both liver lobes. -

Angiocrine Endothelium: from Physiology to Cancer Jennifer Pasquier1,2*, Pegah Ghiabi2, Lotf Chouchane3,4,5, Kais Razzouk1, Shahin Rafi3 and Arash Rafi1,2,3
Pasquier et al. J Transl Med (2020) 18:52 https://doi.org/10.1186/s12967-020-02244-9 Journal of Translational Medicine REVIEW Open Access Angiocrine endothelium: from physiology to cancer Jennifer Pasquier1,2*, Pegah Ghiabi2, Lotf Chouchane3,4,5, Kais Razzouk1, Shahin Rafi3 and Arash Rafi1,2,3 Abstract The concept of cancer as a cell-autonomous disease has been challenged by the wealth of knowledge gathered in the past decades on the importance of tumor microenvironment (TM) in cancer progression and metastasis. The sig- nifcance of endothelial cells (ECs) in this scenario was initially attributed to their role in vasculogenesis and angiogen- esis that is critical for tumor initiation and growth. Nevertheless, the identifcation of endothelial-derived angiocrine factors illustrated an alternative non-angiogenic function of ECs contributing to both physiological and pathological tissue development. Gene expression profling studies have demonstrated distinctive expression patterns in tumor- associated endothelial cells that imply a bilateral crosstalk between tumor and its endothelium. Recently, some of the molecular determinants of this reciprocal interaction have been identifed which are considered as potential targets for developing novel anti-angiocrine therapeutic strategies. Keywords: Angiocrine, Endothelium, Cancer, Cancer microenvironment, Angiogenesis Introduction of blood vessels in initiation of tumor growth and stated Metastatic disease accounts for about 90% of patient that in the absence of such angiogenesis, tumors can- mortality. Te difculty in controlling and eradicating not expand their mass or display a metastatic phenotype metastasis might be related to the heterotypic interaction [7]. Based on this theory, many investigators assumed of tumor and its microenvironment [1]. -

Type of the Paper (Article
Table S1. Gene expression of pro-angiogenic factors in tumor lymph nodes of Ibtk+/+Eµ-myc and Ibtk+/-Eµ-myc mice. Fold p- Symbol Gene change value 0,007 Akt1 Thymoma viral proto-oncogene 1 1,8967 061 0,929 Ang Angiogenin, ribonuclease, RNase A family, 5 1,1159 481 0,000 Angpt1 Angiopoietin 1 4,3916 117 0,461 Angpt2 Angiopoietin 2 0,7478 625 0,258 Anpep Alanyl (membrane) aminopeptidase 1,1015 737 0,000 Bai1 Brain-specific angiogenesis inhibitor 1 4,0927 202 0,001 Ccl11 Chemokine (C-C motif) ligand 11 3,1381 149 0,000 Ccl2 Chemokine (C-C motif) ligand 2 2,8407 298 0,000 Cdh5 Cadherin 5 2,5849 744 0,000 Col18a1 Collagen, type XVIII, alpha 1 3,8568 388 0,003 Col4a3 Collagen, type IV, alpha 3 2,9031 327 0,000 Csf3 Colony stimulating factor 3 (granulocyte) 4,3332 258 0,693 Ctgf Connective tissue growth factor 1,0195 88 0,000 Cxcl1 Chemokine (C-X-C motif) ligand 1 2,67 21 0,067 Cxcl2 Chemokine (C-X-C motif) ligand 2 0,7507 631 0,000 Cxcl5 Chemokine (C-X-C motif) ligand 5 3,921 328 0,000 Edn1 Endothelin 1 3,9931 042 0,001 Efna1 Ephrin A1 1,6449 601 0,002 Efnb2 Ephrin B2 2,8858 042 0,000 Egf Epidermal growth factor 1,726 51 0,000 Eng Endoglin 0,2309 467 0,000 Epas1 Endothelial PAS domain protein 1 2,8421 764 0,000 Ephb4 Eph receptor B4 3,6334 035 V-erb-b2 erythroblastic leukemia viral oncogene homolog 2, 0,000 Erbb2 3,9377 neuro/glioblastoma derived oncogene homolog (avian) 024 0,000 F2 Coagulation factor II 3,8295 239 1 0,000 F3 Coagulation factor III 4,4195 293 0,002 Fgf1 Fibroblast growth factor 1 2,8198 748 0,000 Fgf2 Fibroblast growth factor -

Targeting Fibroblast Growth Factor Pathways in Endometrial Cancer
Curr Probl Cancer 41 (2017) 37–47 Contents lists available at ScienceDirect Curr Probl Cancer journal homepage: www.elsevier.com/locate/cpcancer Targeting fibroblast growth factor pathways in endometrial cancer Boris Winterhoff, MDa, Gottfried E. Konecny, MDb,* article info abstract Novel treatments that improve outcomes for patients with Keywords: recurrent or metastatic endometrial cancer (EC) remain an Endometrial cancer unmet need. Aberrant signaling by fibroblast growth factors Fibroblast growth factor (FGFs) and FGF receptors (FGFRs) has been implicated in Angiogenesis several human cancers. Activating mutations in FGFR2 have been found in up to 16% of ECs, suggesting an opportunity for targeted therapy. This review summarizes the role of the FGF pathway in angiogenesis and EC, and provides an overview of FGFR-targeted therapies under clinical develop- ment for the treatment of EC. & 2017 The Authors. Published by Elsevier Inc. This is an open access article under the CC BY-NC-ND license (http://creativecommons.org/licenses/by-nc-nd/4.0/). Introduction Endometrial cancer (EC) is the most common gynecologic cancer, with 54,870 new cases and 10,170 related deaths estimated for the United States in 2015.1 Localized disease is often curable with surgery and the 5-year relative survival rate for patients diagnosed with early-stage EC is high.1-3 However, for patients who present with metastatic disease or experience a recurrence, prognosis is poor.1-3 Response of metastatic EC to standard therapies (eg, adjuvant radiotherapy, brachytherapy, or chemotherapy) is limited and overall survival for most patients with recurrent or metastatic disease is r1year.2-4 Thus, novel treatment options for this disease remain an unmet need. -

Investigating Hypoxic Tumor Physiology Through Gene Expression Patterns
Oncogene (2003) 22, 5907–5914 & 2003 Nature Publishing Group All rights reserved 0950-9232/03 $25.00 www.nature.com/onc Investigating hypoxic tumor physiology through gene expression patterns Nicholas C Denko*,1, Lucrezia A Fontana1, Karen M Hudson1, Patrick D Sutphin1, Soumya Raychaudhuri2, Russ Altman2 and Amato J Giaccia1 1Division of Radiation and Cancer Biology, Department of Radiation Oncology, Stanford University School of Medicine, Stanford, CA 94305, USA; 2Department of Genetics, Stanford University School of Medicine, Stanford, CA 94305, USA Clinical evidence shows that tumor hypoxia is an relyupon oxygen-dependent energyproduction suffer independent prognostic indicator of poor patient outcome. when oxygen falls below a certain threshold level. Hypoxic tumors have altered physiologic processes, However, too much oxygen can also result in the including increased regions of angiogenesis, increased generation of toxic, damaging radical byproducts. The local invasion, increased distant metastasis and altered net effect of these opposing pressures necessitates apoptotic programs. Since hypoxia is a potent controller the evolution of molecular mechanisms to regulate the of gene expression, identifying hypoxia-regulated genes is cellular availabilityand usage of oxygen. a means to investigate the molecular response to hypoxic The response to low-oxygen conditions has been stress. Traditional experimental approaches have identi- identified in both procaryotes and eucaryotes. Although fied physiologic changes in hypoxic cells. Recent studies the mechanisms for gene regulation in response to low- have identified hypoxia-responsive genes that may define oxygen conditions are diverse, parallel systems have the mechanism(s) underlying these physiologic changes. evolved in different organisms that are transcriptionally For example, the regulation of glycolytic genes by responsive to low-oxygen. -

The Tyrosine-Kinase Inhibitor Sunitinib Targets Vascular Endothelial (VE)-Cadherin: a Marker of Response to Antitumoural Treatment in Metastatic Renal Cell Carcinoma
www.nature.com/bjc ARTICLE Translational Therapeutics The tyrosine-kinase inhibitor sunitinib targets vascular endothelial (VE)-cadherin: a marker of response to antitumoural treatment in metastatic renal cell carcinoma Helena Polena1, Julie Creuzet1, Maeva Dufies2, Adama Sidibé1, Abir Khalil-Mgharbel1, Aude Salomon1, Alban Deroux3, Jean-Louis Quesada4, Caroline Roelants1, Odile Filhol1, Claude Cochet1, Ellen Blanc5, Céline Ferlay-Segura5, Delphine Borchiellini6, Jean-Marc Ferrero6, Bernard Escudier7, Sylvie Négrier5, Gilles Pages8 and Isabelle Vilgrain1 BACKGROUND: Vascular endothelial (VE)-cadherin is an endothelial cell-specific protein responsible for endothelium integrity. Its adhesive properties are regulated by post-translational processing, such as tyrosine phosphorylation at site Y685 in its cytoplasmic domain, and cleavage of its extracellular domain (sVE). In hormone-refractory metastatic breast cancer, we recently demonstrated that sVE levels correlate to poor survival. In the present study, we determine whether kidney cancer therapies had an effect on VE- cadherin structural modifications and their clinical interest to monitor patient outcome. METHODS: The effects of kidney cancer biotherapies were tested on an endothelial monolayer model mimicking the endothelium lining blood vessels and on a homotypic and heterotypic 3D cell model mimicking tumour growth. sVE was quantified by ELISA in renal cell carcinoma patients initiating sunitinib (48 patients) or bevacizumab (83 patients) in the first-line metastatic setting (SUVEGIL and TORAVA trials). RESULTS: Human VE-cadherin is a direct target for sunitinib which inhibits its VEGF-induced phosphorylation and cleavage on endothelial monolayer and endothelial cell migration in the 3D model. The tumour cell environment modulates VE-cadherin functions through MMPs and VEGF. We demonstrate the presence of soluble VE-cadherin in the sera of mRCC patients (n = 131) which level at baseline, is higher than in a healthy donor group (n = 96). -
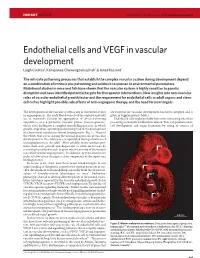
Endothelial Cells and VEGF in Vascular Development Leigh Coultas1, Kallayanee Chawengsaksophak1 & Janet Rossant1
Insight Rossant p7-15 6/12/05 10:23 AM Page 9377 INSIGHT REVIEW NATURE|Vol 438|15 December 2005|doi:10.1038/nature04479 Endothelial cells and VEGF in vascular development Leigh Coultas1, Kallayanee Chawengsaksophak1 & Janet Rossant1 The intricate patterning processes that establish the complex vascular system during development depend on a combination of intrinsic pre-patterning and extrinsic responses to environmental parameters. Mutational studies in mice and fish have shown that the vascular system is highly sensitive to genetic disruption and have identified potential targets for therapeutic interventions. New insights into non-vascular roles of vascular endothelial growth factor and the requirement for endothelial cells in adult organs and stem- cell niches highlight possible side effects of anti-angiogenic therapy and the need for new targets. The development of the vascular system is one of the earliest events are essential for vascular development has been compiled and is in organogenesis. The early blood vessels of the embryo and yolk given in Supplementary Table 1. sac in mammals develop by aggregation of de-novo-forming Endothelial cells might probably have more interesting roles than angioblasts into a primitive vascular plexus (vasculogenesis), just acting as channels for blood circulation. They can promote stem- which then undergoes a complex remodelling process, in which cell development and organ formation by acting as sources of growth, migration, sprouting and pruning lead to the development of a functional circulatory system (angiogenesis; Fig. 1). Many of the events that occur during the normal progression of vascular development in the embryo are recapitulated during situations of neoangiogenesis in the adult1. -

Factor-The Controlling Growth Factor-At a Sufficiently Low Value, While the Cell Population Has Ceased to Grow and Is in a Sort
708 GENETICS: NO VICK A ND SZILARD PROC. N. A. S. EXPERIMENTS WITH THE CHEMOSTAT ON SPONTANEOUS MUTATIONS OF BACTERIA By AARON NOVICK AND LEO SZILARD INSTITUTRE OF RADIOBIOLOGY AND BIOPHYSICS, UNIVERSITY OF CHICAGO Communicated by H. J. Muller, October 18, 1950 Introduction.-All bacteria require for growth the presence of certain inorganic chemical components in the nutrient, such as potassium, phos- phorus, sulphur, etc., and with a few exceptions all bacteria require an energy-yielding carbon source, such as, for instance, glucose or lactate, etc. In addition to these elements or simple compounds, certain bacteria require more complex compounds, for instance an amino acid, which they are not capable of synthesizing. For the purposes of this presentation, any of the chemical compounds which a given strain of bacteria requires for its growth will be called a "growth factor." In general, the growth rate of a bacterial strain may be within very wide limits independent of the concentration of a given growth factor; but since at zero concentration the growth rate is zero, there must of necessity exist, at sufficiently low concentrations of the growth factor, a region in which the growth rate falls with falling concentration of the growth factor. It therefore should be possible to maintain a bacterial population over an indefinite period of time growing at a rate considerably lower than normal simply by maintaining the concentration of one growth factor-the controlling growth factor-at a sufficiently low value, while the concentrations of all other growth factors may at the same time be main- tained at high values.