Migrant Ring Ouzels at a Stopover Site on the South Downs
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

345 Fieldfare Put Your Logo Here
Javier Blasco-Zumeta & Gerd-Michael Heinze Sponsor is needed. Write your name here Put your logo here 345 Fieldfare Fieldfare. Adult. Male (09-I). Song Thrush FIELDFARE (Turdus pilaris ) IDENTIFICATION 25-26 cm. Grey head; red-brown back; grey rump and dark tail; pale underparts; pale flanks spotted black; white underwing coverts; yellow bill with ochre tip. Redwing Fieldfare. Pattern of head, underwing co- verts and flank. SIMILAR SPECIES Song Thrush has orange underwing coverts; Redwing has reddish underwing coverts; Mistle Thrush has white underwing coverts, but lacks pale supercilium and its rump isn’t grey. Mistle Thrush http://blascozumeta.com Write your website here Page 1 Javier Blasco-Zumeta & Gerd-Michael Heinze Sponsor is needed. Write your name here Put your logo here 345 Fieldfare SEXING Male with dark or black tail feathers; red- dish feathers on back with blackish center; most have a broad mark on crown feathers. Female with dark brown tail feathers but not black; dull reddish feathers on back with dark centre (but not blackish); most have a thin mark on crown feathers. CAUTION: some birds of both sexes have similar pattern on crown feathers. Fieldfare. Sexing. Pattern of tail: left male; right fe- male. Fieldfare. Sexing. Pat- AGEING tern of Since this species doesn’t breed in Aragon, only crown feat- 2 age groups can be recognized: hers: top 1st year autumn/2nd year spring with moult male; bot- limit within moulted chestnut inner greater co- tom female. verts and retained juvenile outer greater coverts, shorter and duller with traces of white tips; pointed tail feathers. -

What Is So Rare?
WHAT IS SO RARE? As a Fieldfare in April? In Massachusetts, perhaps only a Western Reef Heron or a White-faced Ibis. April here is a month of avian surprises. The 1986 birdwatching spring was off to a happy start with the discovery by schoolteacher Ralph Richards, during a lonely vigil on the cold and rainy Sunday of April 6, of a Fieldfare Turdus pilaris) in a Concord cornfield near the Sudbury River. According ta a bulletin promptly issued by the Massachusetts Audubon Society, this is a first state record, and there have been, s-ince 1878, ten prior records in North America, the most recent being the occurrence of four Fieldfares in St. John's, Newfoundland, from December 1985 to mid-January 1986. "It seems not unlikely that the Massachusetts bird originated with the same midwinter flight that brought the Newfoundland birds to North America. ..." In winter, the Fieldfare ranges from southern Scandinavia, the British Isles, and central Europe south to the Mediterranean. Outside of the breeding season, the bird's habitat choice is open fields and pastures, where it occurs gregariously in large flocks, often accompanied by Redwings (Turdus iliacus). The Fieldfare nests in northern Scandinavia (it is Norway's most common thrush), in north and central continental Europe east to Siberia, and also in Iceland and southern Greenland. The story of the establishment of the disjunct Greenland population is well documented. In January 1937, a strong southeast gale swept a flock of Fieldfares across Europe and the Atlantic to the island of Jan Mayen and the Greenland coast. -

European Red List of Birds 2015
Turdus torquatus (Ring Ouzel) European Red List of Birds Supplementary Material The European Union (EU27) Red List assessments were based principally on the official data reported by EU Member States to the European Commission under Article 12 of the Birds Directive in 2013-14. For the European Red List assessments, similar data were sourced from BirdLife Partners and other collaborating experts in other European countries and territories. For more information, see BirdLife International (2015). Contents Reported national population sizes and trends p. 2 Trend maps of reported national population data p. 4 Sources of reported national population data p. 6 Species factsheet bibliography p. 10 Recommended citation BirdLife International (2015) European Red List of Birds. Luxembourg: Office for Official Publications of the European Communities. Further information http://www.birdlife.org/datazone/info/euroredlist http://www.birdlife.org/europe-and-central-asia/european-red-list-birds-0 http://www.iucnredlist.org/initiatives/europe http://ec.europa.eu/environment/nature/conservation/species/redlist/ Data requests and feedback To request access to these data in electronic format, provide new information, correct any errors or provide feedback, please email [email protected]. THE IUCN RED LIST OF THREATENED SPECIES™ BirdLife International (2015) European Red List of Birds Turdus torquatus (Ring Ouzel) Table 1. Reported national breeding population size and trends in Europe1. Country (or Population estimate Short-term population trend4 Long-term -

Scottish Birds 22: 9-19
Scottish Birds THE JOURNAL OF THE SOC Vol 22 No 1 June 2001 Roof and ground nesting Eurasian Oystercatchers in Aberdeen The contrasting status of Ring Ouzels in 2 areas of upper Deeside The distribution of Crested Tits in Scotland during the 1990s Western Capercaillie captures in snares Amendments to the Scottish List Scottish List: species and subspecies Breeding biology of Ring Ouzels in Glen Esk Scottish Birds The Journal of the Scottish Ornithologists' Club Editor: Dr S da Prato Assisted by: Dr I Bainbridge, Professor D Jenkins, Dr M Marquiss, Dr J B Nelson, and R Swann Business Editor: The Secretary sac, 21 Regent Terrace Edinburgh EH7 5BT (tel 0131-5566042, fax 0131 5589947, email [email protected]). Scottish Birds, the official journal of the Scottish Ornithologists' Club, publishes original material relating to ornithology in Scotland. Papers and notes should be sent to The Editor, Scottish Birds, 21 Regent Terrace, Edinburgh EH7 SBT. Two issues of Scottish Birds are published each year, in June and in December. Scottish Birds is issued free to members of the Scottish Ornithologists' Club, who also receive the quarterly newsletter Scottish Bird News, the annual Scottish Bird Report and the annual Raplor round up. These are available to Institutions at a subscription rate (1997) of £36. The Scottish Ornithologists' Club was formed in 1936 to encourage all aspects of ornithology in Scotland. It has local branches which meet in Aberdeen, Ayr, the Borders, Dumfries, Dundee, Edinburgh, Glasgow, Inverness, New Galloway, Orkney, St Andrews, Stirling, Stranraer and Thurso, each with its own programme of field meetings and winter lectures. -

Bird Species Recorded in Alvechurch Parish 2010-2016 A) Total in Grid Squares SP0172, SP0272, SP0273, SP0274, SP0275, SP0276, SP
Bird Species Recorded in Alvechurch Parish 2010-2016 A) Total in grid squares SP0172, SP0272, SP0273, SP0274, SP0275, SP0276, SP0370, SP0371, SP0372, SP0374, SP0375, SP0376, SP0469, SP0470, SP0471, SP0472, SP0473, SP0474, SP0475, SP0476, SP0569, SP0570, SP0571, SP0572, SP0573, SP0574, SP0575 Barn Owl Green Sandpiper Pochard Barnacle Goose Green Woodpecker Red Kite Blackbird Greenfinch Redshank Blackcap Grey Heron Redwing Black-headed Gull Grey Wagtail Reed Bunting Blue Tit Greylag Goose Reed Warbler Bullfinch Herring Gull Ring Ouzel Buzzard Hobby Robin Canada Goose House Martin Rook Carrion Crow House Sparrow Sand Martin Caspian Gull Jackdaw Scaup Chaffinch Jay Sedge Warbler Chiffchaff Kestrel Shoveler Coal Tit Kingfisher Siskin Collared Dove Lapwing Skylark Common Gull Lesser Black-backed Gull Smew Common Sandpiper Lesser Redpoll Snipe Common Tern Lesser Whitethroat Song Thrush Coot Linnet Sparrowhawk Cormorant Little Egret Starling Cuckoo Little Grebe Stock Dove Curlew Little Owl Stonechat Dunnock Long-tailed Tit Swallow Feral Pigeon Magpie Swift Fieldfare Mallard Teal Gadwall Mandarin Treecreeper Garden Warbler Meadow Pipit Tufted Duck Goldcrest Mistle Thrush Turnstone Golden Plover Moorhen Wheatear Goldeneye Mute Swan Whitethroat Goldfinch Nuthatch Wigeon Goosander Osprey Willow Warbler Great Crested Grebe Oystercatcher Wood Pigeon Great Grey Shrike Peregrine Woodcock Great Northern Diver Pheasant Wren Great Spotted Woodpecker Pied Wagtail Yellowhammer Great Tit Bird Species Recorded in Alvechurch Parish 2010-2016 B) Individual grid -

EU Business and Biodiversity Platform
EU Business and Biodiversity Platform Workstream 2: Innovation for Biodiversity and Business Innovation for reed bed biomass fuel and biodiversity by FIELDFARE – ANALYSIS OF THE OPPORTUNITY October 2015 Author: Guy Duke Contributing author: Paul Goriup (FIELDFARE) 1 BASIC INFORMATION 1.1 Companies and contact points FIELDFARE Contact: Paul Goriup, Managing Director, FIELDFARE Email: [email protected] Web: www.fieldfare.biz 1.2 Summary of the opportunity An innovation offering significant benefit to small rural businesses in a particular region of the EU (lower Danube) and offering significant enhancement of reed bed biodiversity in this region as well as larger populations along flyways of migrants benefiting from these reed bed enhancements. It offers a financially sustainable solution to reducing the extent of overgrown reed beds and eventually reverse ecologically unsustainable floodplain management in the lower Danube region. Good scalability in the lower Danube region with possible transfer to other reed bed areas in Europe, limited by reed bed area. There is a potential €40+ million per annum market from the lower Danube resource, generating 2000-2500 jobs. Feasibility currently constrained by inflexible standards, slow rate of innovation in management and processing technologies, and low diffusion of appropriate boiler equipment. 1.3 Description of the innovation FIELDFARE harnesses ethical investment in Western Europe to promote ecologically sustainable development and wise use of natural resources in Eastern Europe. It is particularly active in the Danube Delta, a region of high biodiversity, where relatively modest investment can generate large biodiversity benefits. The main innovation is the production of biomass fuel from overgrown, low biodiversity reed beds that have become widespread as a result of floodplain modification since the 1950s. -

Postfledging Survival, Movements, and Dispersal of Ring Ouzels (Turdus Torquatus) Author(S): Innes M
Postfledging Survival, Movements, and Dispersal of Ring Ouzels (Turdus torquatus) Author(s): Innes M. W. Sim , Sonja C. Ludwig , Murray C. Grant , Joanna L. Loughrey , Graham W. Rebecca , and Jane M. Reid Source: The Auk, 130(1):69-77. 2013. Published By: The American Ornithologists' Union URL: http://www.bioone.org/doi/full/10.1525/auk.2012.12008 BioOne (www.bioone.org) is a nonprofit, online aggregation of core research in the biological, ecological, and environmental sciences. BioOne provides a sustainable online platform for over 170 journals and books published by nonprofit societies, associations, museums, institutions, and presses. Your use of this PDF, the BioOne Web site, and all posted and associated content indicates your acceptance of BioOne’s Terms of Use, available at www.bioone.org/page/terms_of_use. Usage of BioOne content is strictly limited to personal, educational, and non-commercial use. Commercial inquiries or rights and permissions requests should be directed to the individual publisher as copyright holder. BioOne sees sustainable scholarly publishing as an inherently collaborative enterprise connecting authors, nonprofit publishers, academic institutions, research libraries, and research funders in the common goal of maximizing access to critical research. The Auk 130(1):69−77, 2013 © The American Ornithologists’ Union, 2013. Printed in USA. POSTFLEDGING SURVIVAL, MOVEMENTS, AND DISPERSAL OF RING OUZELS (TURDUS TORQUATUS) INNES M. W. SIM,1,2,4 SONJA C. LUDWIG,1,5 MURRAY C. GRANT,1,6 JOANNA L. LOUGHREY,2,6 GRAHAM -

Developing Methods for the Field Survey and Monitoring of Breeding Short-Eared Owls (Asio Flammeus) in the UK: Final Report from Pilot Fieldwork in 2006 and 2007
BTO Research Report No. 496 Developing methods for the field survey and monitoring of breeding Short-eared owls (Asio flammeus) in the UK: Final report from pilot fieldwork in 2006 and 2007 A report to Scottish Natural Heritage Ref: 14652 Authors John Calladine, Graeme Garner and Chris Wernham February 2008 BTO Scotland School of Biological and Environmental Sciences, University of Stirling, Stirling, FK9 4LA Registered Charity No. SC039193 ii CONTENTS LIST OF TABLES................................................................................................................... iii LIST OF FIGURES ...................................................................................................................v LIST OF FIGURES ...................................................................................................................v LIST OF APPENDICES...........................................................................................................vi SUMMARY.............................................................................................................................vii EXECUTIVE SUMMARY ................................................................................................... viii CRYNODEB............................................................................................................................xii ACKNOWLEDGEMENTS....................................................................................................xvi 1. BACKGROUND AND AIMS...........................................................................................2 -

Rejection Behavior by Common Cuckoo Hosts Towards Artificial Brood Parasite Eggs
REJECTION BEHAVIOR BY COMMON CUCKOO HOSTS TOWARDS ARTIFICIAL BROOD PARASITE EGGS ARNE MOKSNES, EIVIN ROSKAFT, AND ANDERS T. BRAA Departmentof Zoology,University of Trondheim,N-7055 Dragvoll,Norway ABSTRACT.--Westudied the rejectionbehavior shown by differentNorwegian cuckoo hosts towardsartificial CommonCuckoo (Cuculus canorus) eggs. The hostswith the largestbills were graspejectors, those with medium-sizedbills were mostlypuncture ejectors, while those with the smallestbills generally desertedtheir nestswhen parasitizedexperimentally with an artificial egg. There were a few exceptionsto this general rule. Becausethe Common Cuckooand Brown-headedCowbird (Molothrus ater) lay eggsthat aresimilar in shape,volume, and eggshellthickness, and they parasitizenests of similarly sizedhost species,we support the punctureresistance hypothesis proposed to explain the adaptivevalue (or evolution)of strengthin cowbirdeggs. The primary assumptionand predictionof this hypothesisare that somehosts have bills too small to graspparasitic eggs and thereforemust puncture-eject them,and that smallerhosts do notadopt ejection behavior because of the heavycost involved in puncture-ejectingthe thick-shelledparasitic egg. We comparedour resultswith thosefor North AmericanBrown-headed Cowbird hosts and we found a significantlyhigher propor- tion of rejectersamong CommonCuckoo hosts with graspindices (i.e. bill length x bill breadth)of <200 mm2. Cuckoo hosts ejected parasitic eggs rather than acceptthem as cowbird hostsdid. Amongthe CommonCuckoo hosts, the costof acceptinga parasiticegg probably alwaysexceeds that of rejectionbecause cuckoo nestlings typically eject all hosteggs or nestlingsshortly after they hatch.Received 25 February1990, accepted 23 October1990. THEEGGS of many brood parasiteshave thick- nestseither by grasping the eggs or by punc- er shells than the eggs of other bird speciesof turing the eggs before removal. Rohwer and similar size (Lack 1968,Spaw and Rohwer 1987). -
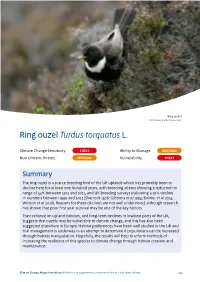
Climate Change Adaptation Manual Second Edition 2020
Ring ouzel © Andy Hay (rspb-images.com) Ring ouzel Turdus torquatus L. Climate Change Sensitivity: HIGH Ability to Manage: MEDIUM Non climatic threats: MEDIUM Vulnerability: HIGH Summary The ring ouzel is a scarce breeding bird of the UK uplands which has probably been in decline here for at least one hundred years, with breeding atlases showing a reduction in range of 44% between 1972 and 2011, and UK breeding surveys indicating a 29% decline in numbers between 1999 and 2012 (Sharrock 1976; Gibbons et al 1993; Balmer et al 2013; Wotton et al 2016). Reasons for these declines are not well understood, although research has shown that poor first year survival may be one of the key factors. Their reliance on upland habitats, and long-term declines in lowland parts of the UK, suggests that ouzels may be vulnerable to climate change, and this has also been suggested elsewhere in Europe. Habitat preferences have been well studied in the UK and trial management is underway in an attempt to determine if populations can be increased through habitat manipulation. Hopefully, the results will help to inform methods of increasing the resilience of this species to climate change through habitat creation and maintenance. Climate Change Adaptation Manual Evidence to support nature conservation in a changing climate 424 Description The ring ouzel is a medium-sized thrush, very similar to but slightly smaller and less stocky than a blackbird, and with slightly longer wings and a distinctive white-cream breast band or gorget in adults. Unlike blackbirds, ouzels have silvery panels in the wings and silver-grey edges to the lower body feathers, often giving them a scaly appearance, which fades with time following the annual moult. -

Population Trends of Common Birds in the Italian Alps
Population Index of Common Breeding Birds in Italian Mountain Prairies Rural development: A critical opportunity for people and biodiversity With a spotlight on the Alpine region Turin 6 november 2013 Mountains cover almost a quarter of the Earth surface, host more than 16% of global human population but provide services to many more communities and people Mountains provide many kind of resources that have positive impacts on human health and its prosperity far beyond their natural boundaries. One for all, water The mountains contribute for the 16% to the Italian national GDP However, despite their importance, the conservation status of mountains is not so satisfactory and many problems exist. Some of these are related with human presence and activities, e.g. effects of climate change, pollution, settlement and infrastructures development and tourism pressure; others with the opposite phenomena e.g. changes in cultural landscape and decrease of open habitats, and their biodiversity, due to the land abandonment For these reasons, MITO2000 project (the Italian national common bird monitoring scheme) decided to focus on species breeding in mountain open habitats MITO2000 is part of PECBMS (Pan-European Common Bird Monitoring Scheme), a Europe-wide network of national breeding bird monitoring projects. This network produces indices of population trends of many breeding bird species. One of the most important index is the FBI (Farmland Bird Index), that is one of the EU indicators to assess the impact of agri- environment measures of the CAP These indices are “aggregated indices” because they are calculated averaging the population trends of some species, choosen on the basis of their shared ecological preferences Through objective procedures we identified a group of species breeding in mountain open habitats and, from 2009, we used them to build up a new aggregate index, named FBIpm ( Mountain Prairies Index) Some results ..... -

4.3 Passerines If You Want to Increase Passerine 1 Birds on Your Moor, This Fact Sheet Helps You Understand Their Habitat and Diet Requirements
BD1228 Determining Environmentally Sustainable and Economically Viable Grazing Systems for the Restoration and Maintenance of Heather Moorland in England and Wales 4.3 Passerines If you want to increase passerine 1 birds on your moor, this fact sheet helps you understand their habitat and diet requirements. The species covered are the commoner moorland passerines that breed in England and Wales: • Meadow pipit • Skylark • Stonechat • Whinchat • Wheatear • Ring ouzel Broad habitat relationships The study examined detailed abundance relationships for the first five species and coarser presence/absence relationships for the last one above. Several other passerine species breed on moorland, from the widespread wren to the rare and highly localised twite, but these were not included in the study. Meadow pipit and skylark occur widely on moorlands, with the ubiquitous meadow pipit being the most abundant moorland bird. Wheatear, whinchat and stonechat are more restricted in where they are found. They tend to be most abundant at lower altitudes and sometimes on relatively steep ground. Wheatears are often associated with old sheepfolds and stone walls that are often used as nesting sites. The increasingly rare ring ouzel is restricted to steep sided valleys and gullies on moorland, often where crags and scree occur. They are found breeding from the lower ground on moorland, up to altitudes of over 800 m. Biodiversity value & status Of the moorland passerine species considered in this study: • Skylark and ring ouzel are red listed in the UK’s Birds of Conservation Concern • Meadow pipit and stonechat are amber listed in the UK’s Birds of Conservation Concern 1 Passerines are songbirds that perch 1 4.3 Passerines • Skylark is on both the England and Welsh Section 74 lists of the Countryside and Rights of Way Act of species the conservation of which will be promoted by the Governments [Note: Skylarks are red listed because of declines on lowland farmland largely, and stonechats are amber listed because of an unfavourable conservation status in Europe.