Rapid Detection of Clostridium Botulinum in Food Using Loop-Mediated Isothermal Amplification (LAMP)
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

The Promising Fuel-Biobutanol
Chapter 6 The Promising Fuel-Biobutanol Hongjuan Liu, Genyu Wang and Jianan Zhang Additional information is available at the end of the chapter http://dx.doi.org/10. 5772/52535 1. Introduction In recent years, two problems roused peoples’ concern. One is energy crisis caused by the depleting of petroleum fuel. The other is environmental issues such as greenhouse effect, global warming, etc. Therefore, renewable sources utilization technology and bioenergy pro‐ duction technology developed fast for solving such two problems. Bioethanol as one of the biofuel has been applied in automobiles with gasoline in different blending proportions (Zhou and Thomson, 2009; Yan and Lin, 2009). Biobutanol is one of the new types of biofuel. It continuously attracted the attention of researchers and industrialists because of its several distinct advantages. 1.1. Property of butanol Butanol is a four carbon straight chained alcohol, colorless and flammable. Butanol can be mixed with ethanol, ether and other organic solvent. Butanol can be used as a solvent, in cosmetics, hydraulic fluids, detergent formulations, drugs, antibiotics, hormones and vita‐ mins, as a chemical intermediate in the production of butyl acrylate and methacrylate, and additionally as an extract agent in the manufacture of pharmaceuticals. Butanol has a 4-car‐ bon structure and the carbon atoms can form either a straight-chain or a branched structure, resulting in different properties. There exist different isomers, based on the location of the– OH and carbon chain structure. The different structures, properties and main applications are shown as Table 1. Although the properties of butanol isomers are different in octane number, boiling point, viscosity, etc., the main applications are similar in some aspects, such as being used as sol‐ vents, industrial cleaners, or gasoline additives. -

Supporting Information
Supporting Information Lozupone et al. 10.1073/pnas.0807339105 SI Methods nococcus, and Eubacterium grouped with members of other Determining the Environmental Distribution of Sequenced Genomes. named genera with high bootstrap support (Fig. 1A). One To obtain information on the lifestyle of the isolate and its reported member of the Bacteroidetes (Bacteroides capillosus) source, we looked at descriptive information from NCBI grouped firmly within the Firmicutes. This taxonomic error was (www.ncbi.nlm.nih.gov/genomes/lproks.cgi) and other related not surprising because gut isolates have often been classified as publications. We also determined which 16S rRNA-based envi- Bacteroides based on an obligate anaerobe, Gram-negative, ronmental surveys of microbial assemblages deposited near- nonsporulating phenotype alone (6, 7). A more recent 16S identical sequences in GenBank. We first downloaded the gbenv rRNA-based analysis of the genus Clostridium defined phylo- files from the NCBI ftp site on December 31, 2007, and used genetically related clusters (4, 5), and these designations were them to create a BLAST database. These files contain GenBank supported in our phylogenetic analysis of the Clostridium species in the HGMI pipeline. We thus designated these Clostridium records for the ENV database, a component of the nonredun- species, along with the species from other named genera that dant nucleotide database (nt) where 16S rRNA environmental cluster with them in bootstrap supported nodes, as being within survey data are deposited. GenBank records for hits with Ͼ98% these clusters. sequence identity over 400 bp to the 16S rRNA sequence of each of the 67 genomes were parsed to get a list of study titles Annotation of GTs and GHs. -

Role of Hydrogen Peroxide Vapor (HPV) for the Disinfection of Hospital Surfaces Contaminated by Multiresistant Bacteria
pathogens Review Role of Hydrogen Peroxide Vapor (HPV) for the Disinfection of Hospital Surfaces Contaminated by Multiresistant Bacteria Michele Totaro, Beatrice Casini , Sara Profeti, Benedetta Tuvo, Gaetano Privitera and Angelo Baggiani * Department of Translational Research and the New Technologies in Medicine and Surgery, University of Pisa, 56123 Pisa, Italy; [email protected] (M.T.); [email protected] (B.C.); [email protected] (S.P.); [email protected] (B.T.); [email protected] (G.P.) * Correspondence: [email protected]; Tel.: 050-221-3583; Fax: 050-221-3588 Received: 10 April 2020; Accepted: 22 May 2020; Published: 24 May 2020 Abstract: The emergence of multiresistant bacterial strains as agents of healthcare-related infection in hospitals has prompted a review of the control techniques, with an added emphasis on preventive measures, namely good clinical practices, antimicrobial stewardship, and appropriate environmental cleaning. The latter item is about the choice of an appropriate disinfectant as a critical role due to the difficulties often encountered in obtaining a complete eradication of environmental contaminations and reservoirs of pathogens. The present review is focused on the effectiveness of hydrogen peroxide vapor, among the new environmental disinfectants that have been adopted. The method is based on a critical review of the available literature on this topic Keywords: hydrogen peroxide vapor; multidrug-resistant bacteria; hospital disinfection 1. Introduction The disinfection of hospital surfaces is a complex operation aimed at reducing the pathogenic microorganism load. An ideal disinfectant must be safe for human health. It may have a good stability in the environment and may be free of toxic activity [1–4]. -

Clostridium Amazonitimonense, Clostridium Me
ORIGINAL ARTICLE Taxonogenomic description of four new Clostridium species isolated from human gut: ‘Clostridium amazonitimonense’, ‘Clostridium merdae’, ‘Clostridium massilidielmoense’ and ‘Clostridium nigeriense’ M. T. Alou1, S. Ndongo1, L. Frégère1, N. Labas1, C. Andrieu1, M. Richez1, C. Couderc1, J.-P. Baudoin1, J. Abrahão2, S. Brah3, A. Diallo1,4, C. Sokhna1,4, N. Cassir1, B. La Scola1, F. Cadoret1 and D. Raoult1,5 1) Aix-Marseille Université, Unité de Recherche sur les Maladies Infectieuses et Tropicales Emergentes, UM63, CNRS 7278, IRD 198, INSERM 1095, Marseille, France, 2) Laboratório de Vírus, Departamento de Microbiologia, Universidade Federal de Minas Gerais, Belo Horizonte, Minas Gerais, Brazil, 3) Hopital National de Niamey, BP 247, Niamey, Niger, 4) Campus Commun UCAD-IRD of Hann, Route des pères Maristes, Hann Maristes, BP 1386, CP 18524, Dakar, Senegal and 5) Special Infectious Agents Unit, King Fahd Medical Research Center, King Abdulaziz University, Jeddah, Saudi Arabia Abstract Culturomics investigates microbial diversity of the human microbiome by combining diversified culture conditions, matrix-assisted laser desorption/ionization time-of-flight mass spectrometry and 16S rRNA gene identification. The present study allowed identification of four putative new Clostridium sensu stricto species: ‘Clostridium amazonitimonense’ strain LF2T, ‘Clostridium massilidielmoense’ strain MT26T, ‘Clostridium nigeriense’ strain Marseille-P2414T and ‘Clostridium merdae’ strain Marseille-P2953T, which we describe using the concept of taxonogenomics. We describe the main characteristics of each bacterium and present their complete genome sequence and annotation. © 2017 Published by Elsevier Ltd. Keywords: ‘Clostridium amazonitimonense’, ‘Clostridium massilidielmoense’, ‘Clostridium merdae’, ‘Clostridium nigeriense’, culturomics, emerging bacteria, human microbiota, taxonogenomics Original Submission: 18 August 2017; Revised Submission: 9 November 2017; Accepted: 16 November 2017 Article published online: 22 November 2017 intestine [1,4–6]. -

Botulism Manual
Preface This report, which updates handbooks issued in 1969, 1973, and 1979, reviews the epidemiology of botulism in the United States since 1899, the problems of clinical and laboratory diagnosis, and the current concepts of treatment. It was written in response to a need for a comprehensive and current working manual for epidemiologists, clinicians, and laboratory workers. We acknowledge the contributions in the preparation of this review of past and present physicians, veterinarians, and staff of the Foodborne and Diarrheal Diseases Branch, Division of Bacterial and Mycotic Diseases (DBMD), National Center for Infectious Diseases (NCID). The excellent review of Drs. K.F. Meyer and B. Eddie, "Fifty Years of Botulism in the United States,"1 is the source of all statistical information for 1899-1949. Data for 1950-1996 are derived from outbreaks reported to CDC. Suggested citation Centers for Disease Control and Prevention: Botulism in the United States, 1899-1996. Handbook for Epidemiologists, Clinicians, and Laboratory Workers, Atlanta, GA. Centers for Disease Control and Prevention, 1998. 1 Meyer KF, Eddie B. Fifty years of botulism in the U.S. and Canada. George Williams Hooper Foundation, University of California, San Francisco, 1950. 1 Dedication This handbook is dedicated to Dr. Charles Hatheway (1932-1998), who served as Chief of the National Botulism Surveillance and Reference Laboratory at CDC from 1975 to 1997. Dr. Hatheway devoted his professional life to the study of botulism; his depth of knowledge and scientific integrity were known worldwide. He was a true humanitarian and served as mentor and friend to countless epidemiologists, research scientists, students, and laboratory workers. -

Detoxification of Lignocellulose-Derived Microbial Inhibitory Compounds by Clostridium Beijerinckii NCIMB 8052 During Acetone-Butanol-Ethanol Fermentation
Detoxification of Lignocellulose-derived Microbial Inhibitory Compounds by Clostridium beijerinckii NCIMB 8052 during Acetone-Butanol-Ethanol Fermentation DISSERTATION Presented in Partial Fulfillment of the Requirements for the Degree Doctor of Philosophy in the Graduate School of The Ohio State University By Yan Zhang Graduate Program in Animal Sciences The Ohio State University 2013 Dissertation Committee: Thaddeus C. Ezeji, Advisor Steven C. Loerch Sandra G. Velleman Zhongtang Yu Venkat Gopalan Copyrighted by Yan Zhang 2013 Abstract Pretreatment and hydrolysis of lignocellulosic biomass to fermentable sugars generate a complex mixture of microbial inhibitors such as furan aldehydes (e.g., furfural), which at sublethal concentration in the fermentation medium can be tolerated or detoxified by acetone butanol ethanol (ABE)-producing Clostridium beijerinckii NCIMB 8052. The response of C. beijerinckii to furfural at the molecular level, however, has not been directly studied. Therefore, this study was to elucidate mechanism employed by C. beijerinckii to detoxify lignocellulose-derived microbial inhibitors and use this information to develop inhibitor-tolerant C. beijerinckii. Towards the long-term goal of developing inhibitor-tolerant Clostridium strains, the first objective was to evaluate ABE fermentation by C. beijerinckii using different proportions of Miscanthus giganteus hydrolysates as carbon source. Compared to the growth of C. beijerinckii in control medium, C. beijerinckii experienced different degrees of inhibition. The degree of inhibition was dose-dependent, and C. beijerinckii did not grow in P2 medium with greater than 25% (v/v) Miscanthus giganteus hydrolysates. To improve tolerance of C. beijerinckii to inhibitors, supplementation of P2 medium with undiluted (100%) Miscanthus giganteus hydrolysates with 4 g/L CaCO3 resulted in successful growth of and ABE production by C. -

Transcriptional Analysis of Degenerate Strain Clostridium Beijerinckii DG
www.nature.com/scientificreports OPEN Transcriptional analysis of degenerate strain Clostridium beijerinckii DG-8052 reveals a Received: 25 August 2016 Accepted: 14 November 2016 pleiotropic response to CaCO3- Published: 14 December 2016 associated recovery of solvent production Shengyin Jiao1,*, Yan Zhang2,*, Caixia Wan3, Jia Lv1, Renjia Du1, Ruijuan Zhang1 & Bei Han1 Degenerate Clostridium beijerinckii strain (DG-8052) can be partially recovered by supplementing CaCO3 to fermentation media. Genome resequencing of DG-8052 showed no general regulator mutated. This study focused on transcriptional analysis of DG-8052 and its response to CaCO3 treatment via microarray. The expressions of 5168 genes capturing 98.6% of C. beijerinckii NCIMB 8052 genome were examed. The results revealed that with addition of CaCO3 565 and 916 genes were significantly up- regulated, and 704 and 1044 genes significantly down-regulated at acidogenic and solventogenic phase of DG-8052, respectively. These genes are primarily responsible for glycolysis to solvent/acid production (poR, pfo), solventogensis (buk, ctf, aldh, adh, bcd) and sporulation (spo0A, sigE, sigma-70, bofA), cell motility and division (ftsA, ftsK, ftsY, ftsH, ftsE, mreB, mreC, mreD, rodA), and molecular chaperones (grpE, dnaK, dnaJ, hsp20, hsp90), etc. The functions of some altered genes in DG-8052, totalling 5.7% at acidogenisis and 8.0% at sovlentogenisis, remain unknown. The response of the degenerate strain to CaCO3 was suggested significantly pleiotropic. This study reveals the multitude of regulatory function that CaCO3 has in clostridia and provides detailed insights into degeneration mechanisms at gene regulation level. It also enables us to develop effective strategies to prevent strain degeneration in future. -
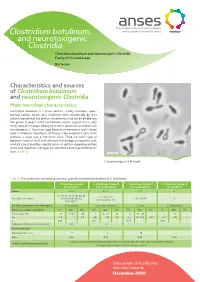
Clostridium Botulinum, and Neurotoxigenic Clostridia Clostridium Botulinum and Neurotoxigenic Clostridia Family of Clostridiaceae Bacterium
Clostridium botulinum, and neurotoxigenic Clostridia Clostridium botulinum and neurotoxigenic Clostridia Family of Clostridiaceae Bacterium Characteristics and sources of Clostridium botulinum and neurotoxigenic Clostridia Main microbial characteristics Clostridium botulinum is a Gram-positive, strictly anaerobic, spore- forming bacillus. Strains of C. botulinum differ considerably by their cultural, biochemical and genetic characteristics and can be divided into four groups (Groups I to IV). Furthermore, certain atypical strains, only rarely isolated in Europe, belonging to other species of Clostridium, are neurotoxigenic: C. butyricum (type E botulinum neurotoxin) and C. baratii (type F botulinum neurotoxin). With only a few exceptions, each strain produces a single type of botulinum toxin. There are seven types of botulinum toxin (A to G) with different immunological properties, each of which is neutralised by a specific serum. In addition, depending on their amino acid sequences, sub-types are identified in each type of botulinum toxin (Table 1). C. botulinum type A. © M. Popoff Table 1. Characteristics concerning survival, growth and toxin production of C. botulinum C. botulinum Group I C. botulinum Group II C. botulinum Group III C. botulinum Group IV (proteolytic) (non-proteolytic) (non-proteolytic) (proteolytic) Toxins A, B, F B, E, F C, D G A1, A2, A3, A4, A5, B1, B2, B3, E1, E2, E3, E6, Sous-types de toxines bivalent B (Ba, Bf, Ab), C, D, C/D, D/C G non-proteolytic B, F proteolytic F Bactéries apparentées non toxinogènes C. sporogenes Pas de nom d’espèces C. novyi C. subterminale Croissance cellules végétatives Min. Opt. Max. Min. Opt. Max. Min. Opt. Max. Min. -

Adaptation and Application of a Two-Plasmid Inducible CRISPR-Cas9 System in Clostridium Beijerinckii
Methods xxx (xxxx) xxx–xxx Contents lists available at ScienceDirect Methods journal homepage: www.elsevier.com/locate/ymeth Adaptation and application of a two-plasmid inducible CRISPR-Cas9 system in Clostridium beijerinckii ⁎ Mamou Dialloa, ,1, Rémi Hocqb,1, Florent Collasa, Gwladys Chartierb, François Waselsb, Hani Surya Wijayaa, Marc W.T. Wertena, Emil J.H. Wolberta, Servé W.M. Kengenc, ⁎ John van der Oostc, Nicolas Lopes Ferreirab, , Ana M. López-Contrerasa a Bioconversion Group, Wageningen Food and Biobased Research, Bornse Weilanden 9, 6708WG Wageningen, the Netherlands b IFP Energies nouvelles, Biotechnology Department, 1 et 4 avenue de Bois-Préau, 92852 Rueil-Malmaison, France c Laboratory of Microbiology, Wageningen University, Stippeneng 4, 6708 WE Wageningen, the Netherlands ARTICLE INFO ABSTRACT Keywords: Recent developments in CRISPR technologies have opened new possibilities for improving genome editing tools Clostridium beijerinckii dedicated to the Clostridium genus. In this study we adapted a two-plasmid tool based on this technology to CRISPR-Cas9 enable scarless modification of the genome of two reference strains of Clostridium beijerinckii producing an Nuclease Acetone/Butanol/Ethanol (ABE) or an Isopropanol/Butanol/Ethanol (IBE) mix of solvents. In the NCIMB 8052 Genome editing ABE-producing strain, inactivation of the SpoIIE sporulation factor encoding gene resulted in sporulation-defi- cient mutants, and this phenotype was reverted by complementing the mutant strain with a functional spoIIE gene. Furthermore, the fungal cellulase-encoding celA gene was inserted into the C. beijerinckii NCIMB 8052 chromosome, resulting in mutants with endoglucanase activity. A similar two-plasmid approach was next used to edit the genome of the natural IBE-producing strain C. -

EXPERIMENTAL STUDIES on FERMENTATIVE FIRMICUTES from ANOXIC ENVIRONMENTS: ISOLATION, EVOLUTION, and THEIR GEOCHEMICAL IMPACTS By
EXPERIMENTAL STUDIES ON FERMENTATIVE FIRMICUTES FROM ANOXIC ENVIRONMENTS: ISOLATION, EVOLUTION, AND THEIR GEOCHEMICAL IMPACTS By JESSICA KEE EUN CHOI A dissertation submitted to the School of Graduate Studies Rutgers, The State University of New Jersey In partial fulfillment of the requirements For the degree of Doctor of Philosophy Graduate Program in Microbial Biology Written under the direction of Nathan Yee And approved by _______________________________________________________ _______________________________________________________ _______________________________________________________ _______________________________________________________ New Brunswick, New Jersey October 2017 ABSTRACT OF THE DISSERTATION Experimental studies on fermentative Firmicutes from anoxic environments: isolation, evolution and their geochemical impacts by JESSICA KEE EUN CHOI Dissertation director: Nathan Yee Fermentative microorganisms from the bacterial phylum Firmicutes are quite ubiquitous in subsurface environments and play an important biogeochemical role. For instance, fermenters have the ability to take complex molecules and break them into simpler compounds that serve as growth substrates for other organisms. The research presented here focuses on two groups of fermentative Firmicutes, one from the genus Clostridium and the other from the class Negativicutes. Clostridium species are well-known fermenters. Laboratory studies done so far have also displayed the capability to reduce Fe(III), yet the mechanism of this activity has not been investigated -

Fimbriae Porphyromonas Gingivalis
TLR2 Transmodulates Monocyte Adhesion and Transmigration via Rac1- and PI3K-Mediated Inside-Out Signaling in Response to Porphyromonas gingivalis This information is current as Fimbriae of September 25, 2021. Evlambia Harokopakis, Mohamad H. Albzreh, Michael H. Martin and George Hajishengallis J Immunol 2006; 176:7645-7656; ; doi: 10.4049/jimmunol.176.12.7645 Downloaded from http://www.jimmunol.org/content/176/12/7645 References This article cites 58 articles, 32 of which you can access for free at: http://www.jimmunol.org/ http://www.jimmunol.org/content/176/12/7645.full#ref-list-1 Why The JI? Submit online. • Rapid Reviews! 30 days* from submission to initial decision • No Triage! Every submission reviewed by practicing scientists by guest on September 25, 2021 • Fast Publication! 4 weeks from acceptance to publication *average Subscription Information about subscribing to The Journal of Immunology is online at: http://jimmunol.org/subscription Permissions Submit copyright permission requests at: http://www.aai.org/About/Publications/JI/copyright.html Email Alerts Receive free email-alerts when new articles cite this article. Sign up at: http://jimmunol.org/alerts The Journal of Immunology is published twice each month by The American Association of Immunologists, Inc., 1451 Rockville Pike, Suite 650, Rockville, MD 20852 Copyright © 2006 by The American Association of Immunologists All rights reserved. Print ISSN: 0022-1767 Online ISSN: 1550-6606. The Journal of Immunology TLR2 Transmodulates Monocyte Adhesion and Transmigration via Rac1- and PI3K-Mediated Inside-Out Signaling in Response to Porphyromonas gingivalis Fimbriae Evlambia Harokopakis,* Mohamad H. Albzreh,‡ Michael H. Martin,*† and George Hajishengallis2*† We present evidence for a novel TLR2 function in transmodulating the adhesive activities of human monocytes in response to the fimbriae of Porphyromonas gingivalis, a pathogen implicated in chronic periodontitis and atherosclerosis. -

Identification and Antimicrobial Susceptibility Testing of Anaerobic
antibiotics Review Identification and Antimicrobial Susceptibility Testing of Anaerobic Bacteria: Rubik’s Cube of Clinical Microbiology? Márió Gajdács 1,*, Gabriella Spengler 1 and Edit Urbán 2 1 Department of Medical Microbiology and Immunobiology, Faculty of Medicine, University of Szeged, 6720 Szeged, Hungary; [email protected] 2 Institute of Clinical Microbiology, Faculty of Medicine, University of Szeged, 6725 Szeged, Hungary; [email protected] * Correspondence: [email protected]; Tel.: +36-62-342-843 Academic Editor: Leonard Amaral Received: 28 September 2017; Accepted: 3 November 2017; Published: 7 November 2017 Abstract: Anaerobic bacteria have pivotal roles in the microbiota of humans and they are significant infectious agents involved in many pathological processes, both in immunocompetent and immunocompromised individuals. Their isolation, cultivation and correct identification differs significantly from the workup of aerobic species, although the use of new technologies (e.g., matrix-assisted laser desorption/ionization time-of-flight mass spectrometry, whole genome sequencing) changed anaerobic diagnostics dramatically. In the past, antimicrobial susceptibility of these microorganisms showed predictable patterns and empirical therapy could be safely administered but recently a steady and clear increase in the resistance for several important drugs (β-lactams, clindamycin) has been observed worldwide. For this reason, antimicrobial susceptibility testing of anaerobic isolates for surveillance