Redalyc.A Temnospondyl Amphibian from the Rio Do Rasto Formation
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Sauropareion Anoplus, with a Discussion of Possible Life History
The postcranial skeleton of the Early Triassic parareptile Sauropareion anoplus, with a discussion of possible life history MARK J. MACDOUGALL, SEAN P. MODESTO, and JENNIFER BOTHA−BRINK MacDougall, M.J., Modesto, S.P., and Botha−Brink, J. 2013. The postcranial skeleton of the Early Triassic parareptile Sauropareion anoplus, with a discussion of possible life history. Acta Palaeontologica Polonica 58 (4): 737–749. The skeletal anatomy of the Early Triassic (Induan) procolophonid reptile Sauropareion anoplus is described on the basis of three partial skeletons from Vangfontein, Middelburg District, South Africa. Together these three specimens preserve the large majority of the pectoral and pelvic girdles, articulated forelimbs and hindlimbs, and all but the caudal portion of the vertebral column, elements hitherto undescribed. Our phylogenetic analysis of the Procolophonoidea is consonant with previous work, positing S. anoplus as the sister taxon to a clade composed of all other procolophonids exclusive of Coletta seca. Previous studies have suggested that procolophonids were burrowers, and this seems to have been the case for S. anoplus, based on comparisons with characteristic skeletal anatomy of living digging animals, such as the presence of a spade−shaped skull, robust phalanges, and large unguals. Key words: Parareptilia, Procolophonidae, phylogenetic analysis, burrowing, Induan, Triassic, South Africa. Mark J. MacDougall [[email protected]], Department of Biology, Cape Breton University, Sydney, Nova Scotia, B1P 6L2, Canada and Department of Biology, University of Toronto at Mississauga, 3359 Mississauga Road, Ontario, L5L 1C6, Canada; Sean P. Modesto [[email protected]], Department of Biology, Cape Breton University, Sydney, Nova Scotia, B1P 6L2, Canada; Jennifer Botha−Brink [[email protected]], Karoo Palaeontology, National Museum, P.O. -

A Procolophonid (Parareptilia) from the Owl Rock Member, Chinle Formation of Utah, Usa
Palaeontologia Electronica http://palaeo-electronica.org A PROCOLOPHONID (PARAREPTILIA) FROM THE OWL ROCK MEMBER, CHINLE FORMATION OF UTAH, USA Nicholas C. Fraser, Randall B. Irmis*, and David K. Elliott ABSTRACT An isolated skull of a procolophonid is described from the Owl Rock Member of the Chinle Formation in the Abajo Mountains of southeast Utah. Although poorly pre- served, this specimen exhibits features that demonstrate a phylogenetic relationship with leptopleuronine procolophonids. These include the dentition, the greatly expanded orbitotemporal opening, the prominent quadratojugal spikes, and the shape of the jugal. Nicholas C. Fraser. Virginia Museum of Natural History, Martinsville, Virginia 24112, USA. [email protected] Randall B. Irmis. Department of Geology, Northern Arizona University, Flagstaff, Arizona 86011, USA. *Current Address: University of California Museum of Paleontology, 1101 Valley Life Sciences Building, Berkeley, California 94720-4780. [email protected] David K. Elliott. Department of Geology, Northern Arizona University, Flagstaff, Arizona 86011, USA. [email protected] KEY WORDS: Procolophonidae; Parareptilia; Late Triassic; Chinle Formation PE Article Number: 8.1.13 Copyright: Society of Vertebrate Paleontology. May 2005 Submission: 28 June 2004. Acceptance: 6 March 2005. INTRODUCTION skull has been described in detail (Kemp 1974; Carroll and Lindsay 1985). The first member of the The Procolophonidae are a group of small clade to be named was Leptopleuron from the parareptiles (sensu Laurin and Reisz -

Alpha Taxonomy of the Russian Permian Procolophonoid Reptiles
Alpha taxonomy of the Russian Permian procolophonoid reptiles LAURA K. SÄILÄ Säilä, L.K. 2009. Alpha taxonomy of the Russian Permian procolophonoid reptiles. Acta Palaeontologica Polonica 54 (4): 599–608. doi:10.4202/app.2009.0017 European Russia has been the source of many procolophonoid taxa from both the Permian and Triassic, and a Permian or− igin for the procolophonoid family Procolophonidae has been based on the Russian taxon Microphon exiguus. Recently, this taxon was reclassified as a seymouriamorph and, in its place, the taxa Nyctiphruretus, Suchonosaurus, and Kinelia from the Middle and Upper Permian of Russia were suggested as “procolophons”, using evolutionary−systematic classifi− cation methods. In recent phylogenies, however, Nyctiphruretus has been recovered as a non–procolophonoid para− reptile, whereas Kinelia and Suchonosaurus have never been included in a phylogenetic study. Re−examination indicates that Suchonosaurus is a member of the procolophonoid subfamily Procolophonidae based on the shape of the maxillary bone and the external naris, the laterally visible maxillary depression, and the number and type of maxillary teeth. Kinelia, on the other hand, is excluded from the Procolophonoidea because of its subpleurodont dental attachment and lack of any procolophonoid features. Thus, Suchonosaurus is the only confirmed Permian procolophonid from the Permian of Rus− sia. Additionally, re−examination of the holotype of Microphon exiguus confirms that it is identical to the seymouria− morph specimens recently included in the genus Microphon and that it lacks procolophonoid features. The earliest un− equivocal record of the subfamily Procolophonidae is confirmed from the Late Permian of Russia, making Russia the only region where, with certainty, both Permian and Triassic procolophonids have been discovered. -

A Reassessment of the Taxonomic Position of Mesosaurs, and a Surprising Phylogeny of Early Amniotes
ORIGINAL RESEARCH published: 02 November 2017 doi: 10.3389/feart.2017.00088 A Reassessment of the Taxonomic Position of Mesosaurs, and a Surprising Phylogeny of Early Amniotes Michel Laurin 1* and Graciela H. Piñeiro 2 1 CR2P (UMR 7207) Centre de Recherche sur la Paléobiodiversité et les Paléoenvironnements (Centre National de la Recherche Scientifique/MNHN/UPMC, Sorbonne Universités), Paris, France, 2 Departamento de Paleontología, Facultad de Ciencias, University of the Republic, Montevideo, Uruguay We reassess the phylogenetic position of mesosaurs by using a data matrix that is updated and slightly expanded from a matrix that the first author published in 1995 with his former thesis advisor. The revised matrix, which incorporates anatomical information published in the last 20 years and observations on several mesosaur specimens (mostly from Uruguay) includes 17 terminal taxa and 129 characters (four more taxa and five more characters than the original matrix from 1995). The new matrix also differs by incorporating more ordered characters (all morphoclines were ordered). Parsimony Edited by: analyses in PAUP 4 using the branch and bound algorithm show that the new matrix Holly Woodward, Oklahoma State University, supports a position of mesosaurs at the very base of Sauropsida, as suggested by the United States first author in 1995. The exclusion of mesosaurs from a less inclusive clade of sauropsids Reviewed by: is supported by a Bremer (Decay) index of 4 and a bootstrap frequency of 66%, both of Michael S. Lee, which suggest that this result is moderately robust. The most parsimonious trees include South Australian Museum, Australia Juliana Sterli, some unexpected results, such as placing the anapsid reptile Paleothyris near the base of Consejo Nacional de Investigaciones diapsids, and all of parareptiles as the sister-group of younginiforms (the most crownward Científicas y Técnicas (CONICET), Argentina diapsids included in the analyses). -
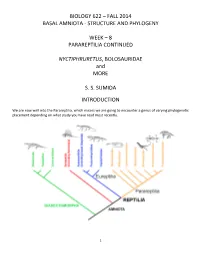
622-2014Lectureweek8
BIOLOGY 622 – FALL 2014 BASAL AMNIOTA - STRUCTURE AND PHYLOGENY WEEK – 8 PARAREPTILIA CONTINUED NYCTIPHRURETUS, BOLOSAURIDAE and MORE S. S. SUMIDA INTRODUCTION We are now well Into the ParareptilIa, whIch means we are goIng to encounter a genus of varyIng phylogenetic placement dependIng on what study you have read most recently. 1 Nyctiphruretus Nyctiphruretus Is a Middle PermIan from the Mezen River BasIn of RussIa that has been varIously classIfIed as a nycteroleterId (part of ParaiesaurIa), a procolophonId, and entirely on Its own. The latter Is the posItion gIven by TsujI and Muller, and that provIsIonally adopted In the phylogeny used for organIzing thIs class. Most recently, SäiIä (2010) placed It as the sIster-taxon to procolophonIds, though that Is still debated. The skull Is generalIzed, wIth fairly sImple teeth suggestive of Insectivory. In other words, It Is prImItive and mIght fIt anywhere. One thIng that Is worth noting is the relative huge orbIts – possIbly suggestive of a nocturnal lIfestyle. 2 THE BOLOSAURIDAE A small, but Important group of [mostly] Early PermIan parareptiles Is the BolosaurIdae. The famIly has been known for over a century, and although Its phylogenetic placement has been all over the map durIng much of that time, membershIp In the BolosaurIdae has been stable and unquestioned. All bolosaurs are characterIzed by dramatic jaw and dental features, particularly the extremely cuspate teeth and the lower jaw wIth extremely hIgh coronoId process. Following are Illustrations of the RussIan form Belebey demonstrating these features: 3 The teeth are unmIstakable, and though somewhat low In dIversIty, bolosaurIds are apparently pan-LaurasIan In dIstrIbution. -

A New Reptile from the Lower Permian of Brazil (Karutia Fortunata Gen
Journal of Systematic Palaeontology ISSN: (Print) (Online) Journal homepage: https://www.tandfonline.com/loi/tjsp20 A new reptile from the lower Permian of Brazil (Karutia fortunata gen. et sp. nov.) and the interrelationships of Parareptilia Juan Carlos Cisneros , Christian F. Kammerer , Kenneth D. Angielczyk , Jörg Fröbisch , Claudia Marsicano , Roger M. H. Smith & Martha Richter To cite this article: Juan Carlos Cisneros , Christian F. Kammerer , Kenneth D. Angielczyk , Jörg Fröbisch , Claudia Marsicano , Roger M. H. Smith & Martha Richter (2021): A new reptile from the lower Permian of Brazil (Karutiafortunata gen. et sp. nov.) and the interrelationships of Parareptilia, Journal of Systematic Palaeontology, DOI: 10.1080/14772019.2020.1863487 To link to this article: https://doi.org/10.1080/14772019.2020.1863487 View supplementary material Published online: 12 Jan 2021. Submit your article to this journal Article views: 107 View related articles View Crossmark data Full Terms & Conditions of access and use can be found at https://www.tandfonline.com/action/journalInformation?journalCode=tjsp20 Journal of Systematic Palaeontology, 2021 http://dx.doi.org/10.1080/14772019.2020.1863487 A new reptile from the lower Permian of Brazil (Karutia fortunata gen. et sp. nov.) and the interrelationships of Parareptilia aà b c b,d Juan Carlos Cisneros , Christian F. Kammerer , Kenneth D. Angielczyk ,Jorg€ Frobisch€ , Claudia Marsicanoe,f , Roger M. H. Smithg,h and Martha Richteri aMuseu de Arqueologia e Paleontologia, Universidade Federal do Piauı, 64049-550 Teresina, Brazil; bPaleontology Unit, North Carolina Museum of Natural Sciences, Raleigh, NC 27601, USA; cNegaunee Integrative Research Center, Field Museum of Natural History, 1400 South Lake Shore Drive, Chicago, IL 60605, USA; dInstitut fur€ Biologie, Humboldt-Universitat€ zu Berlin, Invalidenstr. -

Granja Palmeiras, a New Fossiliferous Site for the Lower Triassic of Southern Brazil
Rev. bras. paleontol. 14(2):157-168, Maio/Agosto 2011 © 2011 by the Sociedade Brasileira de Paleontologia doi:10.4072/rbp.2011.2.04 GRANJA PALMEIRAS, A NEW FOSSILIFEROUS SITE FOR THE LOWER TRIASSIC OF SOUTHERN BRAZIL SÉRGIO DIAS-DA-SILVA Laboratório de Paleobiologia, UNIPAMPA, Campus São Gabriel, Av. Antônio Trilha, 1847, 97300-000, São Gabriel, RS, Brasil. [email protected] ÁTILA AUGUSTO STOCK DA-ROSA Laboratório de Estratigrafia e Paleobiologia, Departamento de Geociências, Centro de Ciências Naturais e Exatas, UFSM, 97105-900, Santa Maria, RS, Brasil. [email protected] ABSTRACT – A new fossiliferous locality, Granja Palmeiras, is described for the Sanga do Cabral Formation (Lower Triassic of Southern Brazil, Rosário do Sul Group, Paraná Basin). It consists on orange and reddish fine sandstones, with sandy and calcareous concretions and intercalated fossiliferous intraformational conglomerates. Its fossils are fragmentary and badly preserved. Nevertheless, among the 46 specimens collected so far, putative actinopterygians, temnospondyls, procolophonoids, archosauromorphs, and non-mammalian cynodonts were identified. Besides, several non-diagnostic appendicular elements are ascribed to Tetrapoda indet. The fossiliferous content found in Granja Palmeiras can provide a valuable new framework for future correlation with other Lower Triassic South American units as well as other continental tetrapod-bearing deposits from Gondwana. Key words: Sanga do Cabral Formation, Lower Triassic, Gondwana, vertebrates. RESUMO – Uma nova localidade fossilífera, Granja Palmeiras, é descrita para a Formação Sanga do Cabral (Triássico Inferior do sul do Brasil, Grupo Rosário do Sul, bacia do Paraná). Consiste em arenitos finos alaranjados e avermelhados, com concreções carbonáticas e arenosas, e conglomerados intraformacionais intercalados. -

Taxonomic Status of the Reptile Genus Procolophon from the Gondwanan Triassic
Taxonomic status of the reptile genus Procolophon from the Gondwanan Triassic Juan Carlos Cisneros* Bernard Price Institute for Palaeontological Research, University of the Witwatersrand, Private Bag 3, WITS, Johannesburg, 2050 South Africa Recieved 29 January 2007. Accepted 29 May 2007 The specific composition of the genus Procolophon in Brazil, South Africa and Antarctica is discussed in the light of new data. It is found that P. pricei and P. brasiliensis, two species described from Brazil, fit within the pattern of ontogenetic variation of the type species P. trigoniceps, and they are here considered junior synonyms. The South African species P. laticeps, characterized by the presence of a temporal fenestra, is no longer considered valid. The peculiar temporal openings of this species are regarded here as an anomalous condition without taxonomic significance. The only complete skull known from Antarctica shows a unique feature consisting of an elliptical depression in the palate. The interpretation of this structure is ambiguous because it may also be attributable to individual variation, and this specimen is provisionally kept within P. trigoniceps. Therefore, only the type species, P. trigoniceps, is recognized in Gondwana. This species occupies a wide geographic range, from the Paraná Basin to the Transantarctic Mountains. Keywords: Procolophonidae, Procolophon, temporal fenestrae, Triassic, Gondwana. INTRODUCTION P. platyrhinus and P. sphenorhinus. Some of the features Procolophon is a member of the Procolophonidae, a clade used to distinguish these species are better explained by of small parareptiles that flourished across all Pangaea taphonomic, ontogenetic or individual variation. Besides, during the Triassic, becoming extinct during the time of all seven of Owen’s and Seeley’s holotypes were collected the T/J boundary. -

Journal of Vertebrate Paleontology a New and Unusual Procolophonid
This article was downloaded by: [Botha-Brink, Jennifer] On: 20 May 2010 Access details: Access Details: [subscription number 922426270] Publisher Taylor & Francis Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37- 41 Mortimer Street, London W1T 3JH, UK Journal of Vertebrate Paleontology Publication details, including instructions for authors and subscription information: http://www.informaworld.com/smpp/title~content=t917000010 A new and unusual procolophonid parareptile from the Lower Triassic Katberg Formation of South Africa Sean P. Modesto a; Diane M. Scott b; Jennifer Botha-Brink cd;Robert R. Reisz b a Department of Biology, Cape Breton University, Sydney, Nova Scotia, Canada b Department of Biology, University of Toronto in Mississauga, Mississauga, Ontario, Canada c Karoo Palaeontology, National Museum, Bloemfontein, South Africa d Department of Zoology and Entomology, University of the Free State, Bloemfontein, South Africa Online publication date: 19 May 2010 To cite this Article Modesto, Sean P. , Scott, Diane M. , Botha-Brink, Jennifer andReisz, Robert R.(2010) 'A new and unusual procolophonid parareptile from the Lower Triassic Katberg Formation of South Africa', Journal of Vertebrate Paleontology, 30: 3, 715 — 723 To link to this Article: DOI: 10.1080/02724631003758003 URL: http://dx.doi.org/10.1080/02724631003758003 PLEASE SCROLL DOWN FOR ARTICLE Full terms and conditions of use: http://www.informaworld.com/terms-and-conditions-of-access.pdf This article may be used for research, teaching and private study purposes. Any substantial or systematic reproduction, re-distribution, re-selling, loan or sub-licensing, systematic supply or distribution in any form to anyone is expressly forbidden. -

The Phylogenetic Position of Nyctiphruretus Acudens, a Parareptile from the Permian of Russia
ISSN (print): 1698-6180. ISSN (online): 1886-7995 www.ucm.es/info/estratig/journal.htm Journal of Iberian Geology 36 (2) 2010: 123-143 doi:10.5209/rev_JIGE.2010.v36.n2.2 The phylogenetic position of Nyctiphruretus acudens, a parareptile from the Permian of Russia Posición filogenética deNyctiphruretus acudens, un parareptil del Pérmico de Rusia L. K. SäiIä1,2 1Department of Earth Sciences, University of Bristol, Wills Memorial Building, Queen’s Road, Bristol, BS8 1JU, United Kingdom 2Department of Geosciences and Geography, P.O. Box 64, Gustaf Hällströmin katu 2a, 00014 University of Helsinki, Finland (current affiliation). [email protected] Received: 09/11/09 / Accepted: 30/12/10 Abstract Several specimens preserving the cranial structure of Nyctiphruretus acudens, a parareptile from the Permian Mezen River locali- ty, Russia, are described here for the first time. Previous studies have offered conflicting reconstructed images ofNyctiphruretus but no illustrations of actual specimens. The new information was incorporated into two existing analyses of parareptilian relationships. The results indicate that Nyctiphruretus is closely related to procolophonoids, pareiasaurs and other non-pareiasaurian Mezen River parareptiles but the interrelationships within this group remain unresolved. Nyctiphruretus was recovered either as the sister taxon of a clade formed of pareiasaurs and the other non-pareiasaurian Mezen River parareptiles, or as the sister taxon of procolophonoids. This is the first time theNyctiphruretus- Procolophonoidea clade received support in a phylogenetic analysis. The interrelationships between several other groups within the Parareptilia also remain unresolved, or poorly supported, highlighting the need for more detailed descriptions and analyses of this enigmatic group of extinct reptiles. -

Barendskraal, a Diverse Amniote Locality from the Lystrosaurus Assemblage Zone, Early Triassic of South Africa
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by Wits Institutional Repository on DSPACE Barendskraal, a diverse amniote locality from the Lystrosaurus Assemblage Zone, Early Triassic of South Africa Ross Damiani1*, Johann Neveling2, Sean Modesto3 & Adam Yates1 1Bernard Price Institute for Palaeontological Research, School of Geosciences, University of the Witwatersrand, Private Bag 3, WITS, 2050 South Africa 2Council for Geoscience, Private Bag X112, Pretoria, 0001 South Africa 3Section of Vertebrate Paleontology, Carnegie Museum of Natural History, 4400 Forbes Avenue, Pittsburgh, Pennsylvania 15213, U.S.A. Received 11 April 2003. Accepted 10 September 2003 A diverse amniote fauna has been recovered from Lower Triassic Lystrosaurus Assemblage Zone exposures on the farm Barendskraal, near Middelburg in Eastern Cape Province, South Africa. The fauna includes the dicynodont therapsid Lystrosaurus sp., the therocephalian therapsids Tetracynodon darti, Moschorhinus kitchingi and Ericiolacerta parva, the archosauromorph reptiles Proterosuchus fergusi and Prolacerta broomi, and the procolophonoid reptiles Owenetta kitchingorum, Sauropareion anoplus and Saurodectes rogersorum. The locality is remarkable in that although it is fossil-rich, Lystrosaurus fossils do not appear to be as abundant as elsewhere in this assem- blage zone, and the diversity of taxa at Barendskraal (at least nine species) is surpassed only by that of the famous Harrismith Common- age locality in the northeastern Free State province (at least 13 species). However, the fauna at Harrismith Commonage is typical of most other Lystrosaurus biozone localities in being dominated numerically by Lystrosaurus. Study of the tetrapod taxa from Barendskraal is providing new insights into procolophonoid phylogeny and survivorship across the Permo-Triassic boundary, as well as the stratigraphic ranges of various taxa in the Lower Triassic deposits of the Karoo Basin. -

A Reevaluation of Early Amniote Phylogeny
Zoological Journal of the Linnean Society (1995), 113: 165–223. With 9 figures A reevaluation of early amniote phylogeny MICHEL LAURIN AND ROBERT R. REISZ* Department of Zoology, Erindale Campus, University of Toronto, Mississauga, Ontario, Canada L5L 1C6 Received February 1994, accepted for publication July 1994 A new phylogenetic analysis of early amniotes based on 124 characters and 13 taxa (including three outgroups) indicates that synapsids are the sister-group of all other known amniotes. The sister-group of Synapsida is Sauropsida, including Mesosauridae and Reptilia as its two main subdivisions. Reptilia is divided into Parareptilia and Eureptilia. Parareptilia includes Testudines and its fossil relatives (Procolophonidae, Pareiasauria and Millerettidae), while Eureptilia includes Diapsida and its fossil relatives (Paleothyris and Captorhinidae). Parts of the phylogeny are robust, such as the sister-group relationship between procolophonids and testudines, and between pareiasaurs and the testudinomorphs (the clade including procolophonids and testudines). Other parts of the new tree are not so firmly established, such as the position of mesosaurs as the sister-group of reptiles. The new phylogeny indicates that three major clades of amniotes extend from the present to the Palaeozoic. These three clades are the Synapsida (including Mammalia), Parareptilia (including Testudines), and Eureptilia (including Sauria). In addition, the Procolophonidae, a group of Triassic parareptiles, are the sister-group of Testudines. ADDITIONAL KEY WORDS:—Amniota – Sauropsida – Mesosauridae – Reptilia – Parareptilia – Eureptilia – Testudines – phylogenetics – evolution – Palaeozoic. CONTENTS Introduction . 166 Methods . 169 Results . 171 Amniote taxonomy . 172 Cotylosauria Cope 1880 . 173 Diadectomorpha Watson 1917 . 173 Amniota Haeckel 1866 . 177 Synapsida Osborn 1903 . 179 Sauropsida Huxley 1864 .