Kinematic Patterns Associated with the Vertical Force Produced During
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Beginner Water Polo Manual
Aquatic Adventures Chapter 6 Water Polo: Splashball and Intro to Water Polo Aquatic Adventures Table of Contents Introduction……………………………………………………….Pg. 3 How to Use this Manual……………………………………..Pg.4 Swimming Skills………………………………………………...Pg. 5 Ball Handling Skills…………………………………………….Pg. 10 Game Skills………………………………………………………...Pg. 16 Game Protocol……………………………………………………Pg. 20 Class Structure and Objective…………………………..Pg. 21 Reference ( Rules; Example Lesson Plans )……………………………….Pg. 23 Bibliography………………………………………………………..Pg. 28 2 Water Polo: Splashball and Intro to Water Polo the most physically fit athletes play due to the nature Water Polo that it is played in water without support from the ground. While this is true, it originated as a far more violent sport with even harder obstacles. The sport’s origins come from northern Europe, specifically the British Isles. Water polo originated as a type of rugby that was played under the water with violence, injuries, and near-drownings as part of the game. The Scots then developed more civilized rules and developed the concept of passing the ball above water while submerging the ball was considered illegal. The sport grew in popularity and spread all across Europe. The Hungarians then took up the sport and developed much of it to the way we play today. They developed the dry pass, meaning that the ball was to never touch the water; this revolutionized the sport and allowed the Hungarians to establish a water polo dynasty. ater polo is one of the oldest Olympic sports. It was included in the 1900 Olympic Games. WOne of the most violent games in history was the 1956 Gold Medal Match between the USSR and Hungary. -

Fina Water Polo Referees Manual 2019-2021
FINA Water Polo Referees’ Manual 2019-2021 FINA WATER POLO REFEREES MANUAL INSTRUCTIONS AND CLARIFICATIONS FOR REFEREES, COACHES and NATIONAL FEDERATIONS. 2019-2021 1 Version: July 2020 FINA Water Polo Referees’ Manual 2019-2021 FINA WORLD LEAGUE 4.2.2 Specific WPWL Rules 4.2.2.1 Field of Play The Field of Play (FOP) for Men shall be 30 x up to 20 metres and for Women 25 x up to 20 metres. In case there is not enough space along the side line to create a lateral substitution area (“flying substitution area”), 1 metre may be deducted from the width of the field of play to create this lateral substitution area. 4.2.2.2 Points Awarded The following points will be awarded per match to each team: Match won: 3 points Match won by penalty: 2 points Match lost by penalty: 1 point Match lost or forfeited: 0 points In the case of a tie in Preliminaries, Semi Finals and Final Games: In all the matches there will be a winner. If at the end of the fourth quarter the score is tied, then the winner shall be resolved by each team shooting five penalty shots from the 5m line in alternate succession. Each team shall select five players to take the penalty shots. The first team shall take its first penalty shot and then the other team shall take its first penalty shot, etc. If a tie shall exist after that procedure, each team shall then take alternate shots until one team scores and the other misses. -

Teams Men Group A
TEAMS MEN GROUP A GERMANY GEORGIA ITALY HUNGARY © LEN MEDIA TEAM EUROPEAN WATER POLO CHAMPIONSHIPS TEAM GERMANY / MEN 2016 Olympic Champion Serbia 2017 World Champion Croatia 2016 European Champion Serbia 2018 World League Winner Montenegro 2014 World Cup Winner Serbia Best results Germany 1928 Olympic champion, 1932 + 1936 silver medallist, 1984 bronze medallist, 1972 and 1988 4th, 2004 5th 1982 World bronze medallist 1981 and 1989 European champion, 2016 European 11th 1985 World Cup champion rd 2005 World League 3 Results DoB 04 SEP 1990 WCh: 2013 10th PoB Duesseldorf ECh: 2014 9th, 2016 11th Residence Duisburg Moritz SCHENKEL Height / Weight 203cm / 103kg Goalkeeper / Righthanded Occupation Student 1 (business administration) Caps: 90 since 2009 Club Waspo 98 Hannover Coach (Club) Karsten Seehafer Active since 2002 Results DoB 27 AUG 1997 PoB Krefeld Ben REIBEL Residence Berlin Attack+Defence+Forward Height / Weight 207cm / 97kg Occupation Student 2 / Righthanded (economic sciences) Club Wasserfreunde Spandau 04 Caps: 15 since 2017 Coach (Club) Petar Kovacevic Active since 2009 Results DoB 29 NOV 1993 ECh: 2014 9th, 2016 11th PoB Heidelberg Timo VAN DER BOSCH Residence Ludwigsburg Height / Weight 194cm / 103kg 3 Center / Righthanded Occupation Police officer Club SV Ludwigsburg 08 Caps: 73 since 2012 Coach (Club) Sven Schulz Active since 2006 Results DoB 22 DEC 1989 Olympics: 2008 10th PoB Oberhausen WCh: 2009 6th, 2011 8th, 2013 10th Julian REAL Residence Hanover ECh: 2012 5th, 2008 and 2010 6th, 2014 9th, CAPTAIN Height / Weight -

Kinematic Analysis of Water Polo Player in the Vertical Thrust
International Journal of Environmental Research and Public Health Article Kinematic Analysis of Water Polo Player in the Vertical Thrust Performance to Determine the Force-Velocity and Power-Velocity Relationships in Water: A Preliminary Study Giuseppe Annino 1,2 , Cristian Romagnoli 2,3 , Andrea Zanela 4, Giovanni Melchiorri 1,5, Valerio Viero 5, Elvira Padua 6,* and Vincenzo Bonaiuto 2 1 Department of Medicine Systems, “Tor Vergata” University of Rome, via Montpellier 1, 00133 Rome, Italy; [email protected] (G.A.); [email protected] (G.M.) 2 Sport Engineering Lab, Department of Industrial Engineering, “Tor Vergata” University of Rome, via del Politecnico 1, 00133 Rome, Italy; [email protected] (C.R.); [email protected] (V.B.) 3 Department for Life Quality Studies, University of Bologna, 47900 Rimini, Italy 4 Robotics and Artificial Intelligence Lab, ENEA “Casaccia” Research Centre, via Anguillarese, 00301 Rome, Italy; [email protected] 5 Italian Swimming Federation, Stadio Olimpico Curva Nord, 00135 Rome, Italy; [email protected] 6 Department of Human Science and Promotion of Quality of Life, San Raffaele Open University of Rome, via di val Cannuta 247, 00166 Rome, Italy * Correspondence: [email protected] Abstract: Background: To date, studies on muscle force and power-velocity (F-v and P-v) relation- Citation: Annino, G.; Romagnoli, C.; ships performed in water are absent. Aim: The goal of this study is to derive the F-v and P-v Zanela, A.; Melchiorri, G.; Viero, V.; regression models of water polo players in water vertical thrust performance at increasing load. Padua, E.; Bonaiuto, V. -

Adam Krikorian– Head Coach • 9Th Year • UCLA ('97)
Adam Krikorian – Head Coach • 9th Year • UCLA (‘97) There may not be another head coach in any a third-place finish in the MPSF Tournament and sport throughout the country who has accomplished a No. 3 final national ranking. For the first time in more than Adam Krikorian in such a short span. In program history, the Bruins won four games against his 16 years with UCLA’s water polo program as conference-rival Stanford. In the spring of 2007, both a player and a coach, Krikorian has won an Krikorian guided the women’s team to its third unprecedented 13 national titles – nine as a head consecutive NCAA title and UCLA’s 100th NCAA coach, three as an assistant coach and one as a team championship. The 2007 women’s water polo student-athlete. title marked Krikorian’s fourth national championship This fall, Adam Krikorian enters his ninth season since the start of the 2004-05 school year. as head coach of the UCLA men’s water polo team That season, Krikorian led both water polo teams and his seventh season alone at the helm. In 1999 to national-champion status for the third time in and 2000, he shared head coaching duties with his head coaching career. Krikorian previously led Guy Baker, who now serves as head coach of the both squads to national championships in the same U.S. Women’s National Team. season in 1999-2000 and 2000-01. The 2004 men’s As the men’s water polo head coach, Krikorian water polo team finished with the best winning per- has guided UCLA to three NCAA Championships centage of any UCLA water polo team since 1972, and boasts a .767 winning percentage (155-47 and the 2005 women’s team completed the second record). -

Muscle Activity During the Typical Water Polo Eggbeater Kick
International Journal of Aquatic Research and Education Volume 4 Number 2 Article 7 5-1-2010 Muscle Activity During the Typical Water Polo Eggbeater Kick Nuno Oliveira University of Porto, Faculty of Sport, [email protected] Ricardo Jorge Fernandes University of Porto, Faculty of Sport Mariana Sarmento University of Porto, Faculty of Sport Silverio Liberal University of Porto, Faculty of Sport Pedro Alexandre Figueiredo University of Porto, Faculty of Sport See next page for additional authors Follow this and additional works at: https://scholarworks.bgsu.edu/ijare Recommended Citation Oliveira, Nuno; Fernandes, Ricardo Jorge; Sarmento, Mariana; Liberal, Silverio; Figueiredo, Pedro Alexandre; Gonçalves, Pedro; and Vilas-Boas, João Paulo (2010) "Muscle Activity During the Typical Water Polo Eggbeater Kick," International Journal of Aquatic Research and Education: Vol. 4 : No. 2 , Article 7. DOI: https://doi.org/10.25035/ijare.04.02.07 Available at: https://scholarworks.bgsu.edu/ijare/vol4/iss2/7 This Research Article is brought to you for free and open access by the Journals at ScholarWorks@BGSU. It has been accepted for inclusion in International Journal of Aquatic Research and Education by an authorized editor of ScholarWorks@BGSU. Muscle Activity During the Typical Water Polo Eggbeater Kick Authors Nuno Oliveira, Ricardo Jorge Fernandes, Mariana Sarmento, Silverio Liberal, Pedro Alexandre Figueiredo, Pedro Gonçalves, and João Paulo Vilas-Boas This research article is available in International Journal of Aquatic Research and Education: https://scholarworks.bgsu.edu/ijare/vol4/iss2/7 Oliveira et al.: Muscle Activity During the Typical Water Polo Eggbeater Kick International Journal of Aquatic Research and Education, 2010, 4, 163-174 © 2010 Human Kinetics, Inc. -

Canoe Polo About the Game
CANOE POLO ABOUT THE GAME INTRODUCTION Five players per team play a game similar to water polo, with a few minor alterations. Each of the 5-aside team, (plus subs) are in kayaks, and the goals are suspended in the air. Canoe polo is similar to basketball in that it is based on possession, passing, blocking, and scoring, though with the addition of tackling under tightly prescribed rules. A team game for all ages where skills will dominate strength, that is extremely safe due to protective equipment, the indoor pool environment, and close adult supervision. GEAR One kayak, one double-bladed paddle, one helmet with facemask, a spray deck and a buoyancy vest. (All provided by WCPA) SOME OF THE RULES NUMBER OF PLAYERS Each team may consist of a maximum of eight (8) players for any one game. No more than five (5) players are permitted on the playing area at any one time. Any other players at that moment are to be considered as substitutes. A team must begin each game with five (5) players, ready to start on or behind their own goal line. If a team is reduced to two (2) players at any time the referee shall end the game. PLAYING TIME Playing time shall be two (2) periods each of eight (8) - ten (10) minutes. The half time interval will be two (2) minutes. The teams shall change ends at half time. START OF GAME At the beginning of each game, five (5) players will line up ready to start, stationary and with some part of their kayaks on or behind their own goal line. -

Water Polo Team to Competitive Level: Chapter 1 Page 1 of 1
Taking a Young Inexperienced Water Polo Team to Competitive Level: Chapter 1 Page 1 of 1 WATER POLO Learning and Teaching the Basics CHAPTER ONE Taking a Young Inexperienced Monte Nitzkowski Water Polo Team to Competitive Level I'm often asked, "Coach, what's the quickest way to become competitive in this sport? We have a new program with young players, and we're getting clobbered." There are four basic axioms which must be followed if you wish to make young teams quickly competitive: CONDITIONING : Get your players into top physical condition. This is the easiest of the four axioms to achieve. Most of the athletes probably have a swimming background and are already in pretty good shape. Hit them hard with a conditioning program which features considerable basic swim training, a lot of ball conditioning and conditioning for quickness. You might not get them in better shape than your opponents, but "equal shape" is good enough. BALLHANDLING: Spend a lot of time on ballhandling. All players need to continue to develop their ballhandling skills—for young players it's a must. Passing is the name of the game. If a player can't pick up the ball and pass it accurately, his/her team will be in constant trouble. Ball control is essential and without it, success is impossible to achieve. Along with basic ballhandling skills, at the earliest stage of development, young players must be taught how to draw a foul. Most often, opponents will press against inexperienced teams. If players can't draw a foul and pass from the free throw, they will find it extremely difficult to move the ball into scoring position. -

INNERTUBE WATER POLO RULES National Intramural-Recreational Sports Association Rules Will Be Used
INNERTUBE WATER POLO RULES National Intramural-Recreational Sports Association rules will be used. Italic print indicates special rules for UNC-Chapel Hill Intramural and/or Co-Recreational play only. Each player is responsible for presenting a current UNC ONE CARD or valid government issued ID at game time. TEAM REPRESENTATIVES Only the captain may speak to the officials. The captain is responsible for notifying his/her team of all the rules as well as anything discussed in the captain’s meeting. THE GAME The game shall be played between two teams of four [4] players. Three players are required to avoid a forfeit. The Co-Recreational game shall be played between two teams of 4 players; two men and two women. Three players are required to avoid a forfeit PENALTY AREA There is an imaginary 2-yard line that extends the entire width of the playing area surrounding the goalkeeper. No part of the offensive player’s body may penetrate the goalkeepers’ area. THE GOALKEEPER Goalkeeper position: 1. The goalkeeper may not throw the ball more than half the length of the pool at any time. 2. The goalkeeper must sit inside the inner tube; the lower half must be inside the inner tube at all times. 3. The goalie may not hold the ball for more than five seconds. 4. Any defensive player may act as the goalkeeper, but there may only be one defensive player inside the goalkeepers’ area at a time. 5. A team may pull their goalie during the last 2 minutes of the game. 6. -
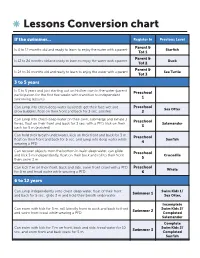
Lessons Conversion Chart
Lessons Conversion chart If the swimmer... Register In Previous Level Parent & Is 4 to 12 months old and ready to learn to enjoy the water with a parent Starfish Tot 1 Parent & Is 12 to 24 months old and ready to learn to enjoy the water with a parent Duck Tot 2 Parent & Is 24 to 36 months old and ready to learn to enjoy the water with a parent Sea Turtle Tot 3 3 to 5 years Is 3 to 5 years and just starting out on his/her own in the water (parent Preschool participation for the first five weeks with transition to independent swimming lessons) 1 Can jump into chest-deep water (assisted), get their face wet and Preschool Sea Otter blow bubbles, float on their front and back for 3 sec. assisted 2 Can jump into chest-deep water on their own, submerge and exhale 3 Preschool times, float on their front and back for 3 sec. with a PFD, kick on their Salamander back for 5 m (assisted) 3 Can hold their breath underwater, kick on their front and back for 3 m, Preschool float on their front and back for 5 sec. and jump into deep water while Sunfish wearing a PFD 4 Can recover objects from the bottom in waist deep water, can glide Preschool and kick 5 m independently, float on their back and roll to their front Crocodile then swim 3 m 5 Can kick 7 m on their front, back and side, swim front crawl with a PFD Preschool Whale for 5 m and tread water while wearing a PFD 6 6 to 12 years Can jump independently unto chest-deep water, float of their front Swim Kids 1/ Swimmer 1 and back for 5 sec., glide 3 m and hold their breath underwater Sea Otter, Incomplete Can swim with kick for 5 m, roll laterally front to back and back to front Swim Kids 2/ Swimmer 2 and swim front crawl while wearing a PFD Completed Salamander Complete Can swim with kick for 7 m on front, back and side, tread water for 10 Swim Kids 2/ Swimmer 3 sec. -

Water Polo for Players and Teachers of Aquatics
1 WATER POLO FOR PLAYERS & TEACHERS OF AQUATICS Pete Snyder, Ph. D. Professor, Fullerton College, Fullerton, California Layout Design: Mary Jo Reutter ©2008, updated 2017. All rights reserved. This manual may not, in whole or in part, be copied, photocopied, reproduced, translated, or converted to any electronic or machine readable form without prior written consent of the author. It is Web-pub- lished by the LA84 Foundation under a license from the author. Printed in the USA 2 Acknowledgements This book would not have been completed and had its particular attention to detail without the help and support of the following individuals. First I’d like to thank Mr. Lundy Smith, an English teacher and head Girls Water Polo Coach at Phillips Exeter Academy in Exeter, New Hampshire. Lundy was most helpful in points of emphasis and grammatical structure in the book. I’d also like to thank Roger Nekton from Phillips Exeter. Roger just retired as head boys water polo and swimming coach after a long and very distinguished career of over thirty years at the Academy. Roger provided valuable technical input as well as encouragement regarding the need for written material in the sport of water polo. As a picture is worth a thousand words, I’m very indebted to Mrs. Chris Kittredge of CMK Enterprises (www.tudorgraphics.com). Chris was able to capture some of the more difficult team aspects of the game only because of her dedication, extensive knowledge of over twenty years with the sport and her brilliance as a photographer. Matt Brown (www.mattbrownphoto.com) is another photographer who exhibited a tremendous amount of alacrity in his water polo photos. -

Hockey Substitute Goalkeeper Guidelines
RETURN TO HOCKEY SUBSTITUTE GOALKEEPER GUIDELINES As a result of the ongoing Covid-19 pandemic, Teams are not able to access affiliate players during the 2020-2021 season. The following guidelines have been established to provide direction to teams on how to manage a situation when their registered goalie(s) are unavailable. Within the Divisions of Senior and Junior, as well as the AAA and AA categories of U18, U16, U15 and U13 the following process(es) will be in place: • When a team only has one goaltender available for a scheduled League game and is seeking access to a back-up goaltender: 1. Teams should consult with each other to determine whether or not the opposing Team will have a properly registered back-up goaltender available. If so, that goaltender should be used as a Substitute should either Team’s goaltender become injured or ill; or 2. If each Team only has 1 properly registered goaltender available for a game, Teams have the ability to apply for access to a Substitute Goalkeeper through the normal application process outlined within the Regulations. ▪ Only one Substitute Goaltender will be considered for a pair/ group of Teams within a ‘Competition Cohort’; ▪ If approved, the Team(s) must ensure that all protocols for using a Substitute Goalkeeper (as outlined below) are adhered to; 3. Or, If an eligible Substitute Goalkeeper is not able to be found or approved, and a goaltender becomes injured or ill, Teams have the ability to dress a player or play the remainder of the game with 6 skaters (as per the Hockey Canada Playing Rules).