Ligase 1 Is a Predictor of Platinum Resistance and Its Blockade Is Synthetically Lethal In
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

LIG1 Antibody Cat
LIG1 Antibody Cat. No.: 26-847 LIG1 Antibody Specifications HOST SPECIES: Rabbit SPECIES REACTIVITY: Human, Mouse Antibody produced in rabbits immunized with a synthetic peptide corresponding a region IMMUNOGEN: of human LIG1. TESTED APPLICATIONS: ELISA, WB LIG1 antibody can be used for detection of LIG1 by ELISA at 1:62500. LIG1 antibody can be APPLICATIONS: used for detection of LIG1 by western blot at 1 μg/mL, and HRP conjugated secondary antibody should be diluted 1:50,000 - 100,000. POSITIVE CONTROL: 1) 721_B Cell Lysate PREDICTED MOLECULAR 102 kDa WEIGHT: Properties PURIFICATION: Antibody is purified by peptide affinity chromatography method. CLONALITY: Polyclonal CONJUGATE: Unconjugated PHYSICAL STATE: Liquid September 29, 2021 1 https://www.prosci-inc.com/lig1-antibody-26-847.html Purified antibody supplied in 1x PBS buffer with 0.09% (w/v) sodium azide and 2% BUFFER: sucrose. CONCENTRATION: batch dependent For short periods of storage (days) store at 4˚C. For longer periods of storage, store LIG1 STORAGE CONDITIONS: antibody at -20˚C. As with any antibody avoid repeat freeze-thaw cycles. Additional Info OFFICIAL SYMBOL: LIG1 ALTERNATE NAMES: LIG1, MGC117397, MGC130025, ACCESSION NO.: NP_000225 PROTEIN GI NO.: 4557719 GENE ID: 3978 USER NOTE: Optimal dilutions for each application to be determined by the researcher. Background and References LIG1is DNA ligase I, with functions in DNA replication and the base excision repair process. Mutations in LIG1 that lead to DNA ligase I deficiency result in immunodeficiency and increased sensitivity to DNA-damaging agents.LIG1 encodes DNA ligase I, with functions in DNA replication and the base excision repair process. -

Regulates Cellular Telomerase Activity by Methylation of TERT Promoter
www.impactjournals.com/oncotarget/ Oncotarget, 2017, Vol. 8, (No. 5), pp: 7977-7988 Research Paper Tianshengyuan-1 (TSY-1) regulates cellular Telomerase activity by methylation of TERT promoter Weibo Yu1, Xiaotian Qin2, Yusheng Jin1, Yawei Li2, Chintda Santiskulvong3, Victor Vu1, Gang Zeng4,5, Zuofeng Zhang6, Michelle Chow1, Jianyu Rao1,5 1Department of Pathology and Laboratory Medicine, David Geffen School of Medicine, University of California at Los Angeles, Los Angeles, CA, USA 2Beijing Boyuantaihe Biological Technology Co., Ltd., Beijing, China 3Genomics Core, Cedars-Sinai Medical Center, Los Angeles, CA, USA 4Department of Urology, David Geffen School of Medicine, University of California at Los Angeles, Los Angeles, CA, USA 5Jonsson Comprehensive Cancer Center, University of California at Los Angeles, Los Angeles, CA, USA 6Department of Epidemiology, School of Public Health, University of California at Los Angeles, Los Angeles, CA, USA Correspondence to: Jianyu Rao, email: [email protected] Keywords: TSY-1, hematopoietic cells, Telomerase, TERT, methylation Received: September 08, 2016 Accepted: November 24, 2016 Published: December 15, 2016 ABSTRACT Telomere and Telomerase have recently been explored as anti-aging and anti- cancer drug targets with only limited success. Previously we showed that the Chinese herbal medicine Tianshengyuan-1 (TSY-1), an agent used to treat bone marrow deficiency, has a profound effect on stimulating Telomerase activity in hematopoietic cells. Here, the mechanism of TSY-1 on cellular Telomerase activity was further investigated using HL60, a promyelocytic leukemia cell line, normal peripheral blood mononuclear cells, and CD34+ hematopoietic stem cells derived from umbilical cord blood. TSY-1 increases Telomerase activity in normal peripheral blood mononuclear cells and CD34+ hematopoietic stem cells with innately low Telomerase activity but decreases Telomerase activity in HL60 cells with high intrinsic Telomerase activity, both in a dose-response manner. -

Iron Depletion Reduces Abce1 Transcripts While Inducing The
Preprints (www.preprints.org) | NOT PEER-REVIEWED | Posted: 22 October 2019 doi:10.20944/preprints201910.0252.v1 1 Research Article 2 Iron depletion Reduces Abce1 Transcripts While 3 Inducing the Mitophagy Factors Pink1 and Parkin 4 Jana Key 1,2, Nesli Ece Sen 1, Aleksandar Arsovic 1, Stella Krämer 1, Robert Hülse 1, Suzana 5 Gispert-Sanchez 1 and Georg Auburger 1,* 6 1 Experimental Neurology, Goethe University Medical School, 60590 Frankfurt am Main; 7 2 Faculty of Biosciences, Goethe-University Frankfurt am Main, Germany 8 * Correspondence: [email protected] 9 10 Abstract: Lifespan extension was recently achieved in Caenorhabditis elegans nematodes by 11 mitochondrial stress and mitophagy, triggered via iron depletion. Conversely in man, deficient 12 mitophagy due to Pink1/Parkin mutations triggers iron accumulation in patient brain and limits 13 survival. We now aimed to identify murine fibroblast factors, which adapt their mRNA expression 14 to acute iron manipulation, relate to mitochondrial dysfunction and may influence survival. After 15 iron depletion, expression of the plasma membrane receptor Tfrc with its activator Ireb2, the 16 mitochondrial membrane transporter Abcb10, the heme-release factor Pgrmc1, the heme- 17 degradation enzyme Hmox1, the heme-binding cholesterol metabolizer Cyp46a1, as well as the 18 mitophagy regulators Pink1 and Parkin showed a negative correlation to iron levels. After iron 19 overload, these factors did not change expression. Conversely, a positive correlation of mRNA levels 20 with both conditions of iron availability was observed for the endosomal factors Slc11a2 and Steap2, 21 as well as for the iron-sulfur-cluster (ISC)-containing factors Ppat, Bdh2 and Nthl1. -

A New Class of Disordered Elements Controls DNA Replication Through
RESEARCH ARTICLE A new class of disordered elements controls DNA replication through initiator self-assembly Matthew W Parker1,2, Maren Bell2, Mustafa Mir2, Jonchee A Kao2, Xavier Darzacq2, Michael R Botchan2*, James M Berger1* 1Department of Biophysics and Biophysical Chemistry, Johns Hopkins School of Medicine, Baltimore, United States; 2Department of Molecular and Cell Biology, University of California, Berkeley, Berkeley, United States Abstract The initiation of DNA replication in metazoans occurs at thousands of chromosomal sites known as origins. At each origin, the Origin Recognition Complex (ORC), Cdc6, and Cdt1 co- assemble to load the Mcm2-7 replicative helicase onto chromatin. Current replication models envisage a linear arrangement of isolated origins functioning autonomously; the extent of inter- origin organization and communication is unknown. Here, we report that the replication initiation machinery of D. melanogaster unexpectedly undergoes liquid-liquid phase separation (LLPS) upon binding DNA in vitro. We find that ORC, Cdc6, and Cdt1 contain intrinsically disordered regions (IDRs) that drive LLPS and constitute a new class of phase separating elements. Initiator IDRs are shown to regulate multiple functions, including chromosome recruitment, initiator-specific co- assembly, and Mcm2-7 loading. These data help explain how CDK activity controls replication initiation and suggest that replication programs are subject to higher-order levels of inter-origin organization. DOI: https://doi.org/10.7554/eLife.48562.001 *For -

Kinetic Analysis of Human DNA Ligase III by Justin R. Mcnally A
Kinetic Analysis of Human DNA Ligase III by Justin R. McNally A dissertation submitted in partial fulfillment of the requirements for the degree of Doctor of Philosophy (Biological Chemistry) in the University of Michigan 2019 Doctoral Committee: Associate Professor Patrick J. O’Brien, Chair Associate Professor Bruce A. Palfey Associate Professor JoAnn M. Sekiguchi Associate Professor Raymond C. Trievel Professor Thomas E. Wilson Justin R. McNally [email protected] ORCID iD: 0000-0003-2694-2410 © Justin R. McNally 2019 Table of Contents List of Tables iii List of Figures iv Abstract vii Chapter 1 Introduction to the human DNA ligases 1 Chapter 2 Kinetic Analyses of Single-Strand Break Repair by Human DNA Ligase III Isoforms Reveal Biochemical Differences from DNA Ligase I 20 Chapter 3 The LIG3 N-terminus, in its entirety, contributes to single-strand DNA break ligation 56 Chapter 4 Comparative end-joining by human DNA ligases I and III 82 Chapter 5 A real-time DNA ligase assay suitable for high throughput screening 113 Chapter 6 Conclusions and Future Directions 137 ii List of Tables Table 2.1: Comparison of kinetic parameters for multiple turnover ligation by human DNA ligases 31 Table 2.2: Comparison of single-turnover parameters of LIG3β and LIG1 34 Table 3.1: Comparison of LIG3β N-terminal mutant kinetic parameters 67 Table 4.1: Rate constants for sequential ligation by LIG3β 95 Table 5.1: Comparison of multiple turnover kinetic parameters determined by real-time fluorescence assay and reported values 129 iii List of Figures Figure -
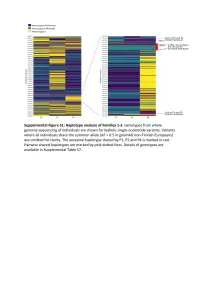
Supplementary Text and Figures.Pdf
Supplemental Figure S2. PRIM1 immunoblots (20-75 kDa). (A) Corresponds to Figure 2D, with additional bands seen in P2 at 75 kDa and 25 kDa. (B) Independent experiment with cell lysates from C and P2 in which additional bands at 25 kDa not evident. (C) Corresponds to Figure 2D. No additional band visible at 25 kDa. (D) Validation of monoclonal antibody (8G10) with siRNA to PRIM1 in HeLa cells. siLUC, luciferase negative control. siPRIM1, siRNA targeting PRIM1 transcript. Vinculin used as a loading control. Supplemental Figure S3: Predicted destabilisation of PRIM1 by the C301R substitution. (A) Cysteine 301 lies in a buried hydrophobic region in PRIM1. DNA primase dimer crystal structure (PDB: 4BPU). PRIM1 residues shaded according to solvent accessibility. Cysteine 301, substituted to arginine in P5, lies in a buried hydrophobic region. (B) Cys301 is evolutionarily conserved in vertebrates. Neither arginine or other large or charged amino acids are observed at this position in other species. (C) The C301R substitution observed in P5 is predicted to lead to destabilisation of all available PRIM1 crystal structures. In contrast, substitution with leucine or threonine, as observed in some orthologous proteins, is not predicted to lead to destabilisation. Data points, predicted changes in free-energy (ΔΔG) from FoldX plotted for each of 14 available crystal structures. (D) Immunoblotting of RPE1 cells transfected with dual reporter constructs (as described in Figure 3B and Materials and Methods) confirms production of PRIM1-GFP and FLAG-SR at expected molecular weights and shows reduced protein levels for the C301R variant. n=2 experiments shown. C301R protein levels relative to WT after normalization to actin loading control indicated underneath the blot. -

Tricarboxylic Acid Cycle Metabolites As Mediators of DNA Methylation Reprogramming in Bovine Preimplantation Embryos
Supplementary Materials Tricarboxylic Acid Cycle Metabolites as Mediators of DNA Methylation Reprogramming in Bovine Preimplantation Embryos Figure S1. (A) Total number of cells in fast (FBL) and slow (SBL) blastocysts; (B) Fluorescence intensity for 5-methylcytosine and 5-hydroxymethylcytosine of fast and slow blastocysts of cells from Trophoectoderm (TE) or inner cell mass (ICM). Fluorescence intensity for 5-methylcytosine of cells from the ICM or TE in blastocysts cultured with (C) dimethyl-succinate or (D) dimethyl-α- ketoglutarate. Statistical significance is identified by different letters. Figure S2. Experimental design. Table S1. Selected genes related to metabolism and epigenetic mechanisms from RNA-Seq analysis of bovine blastocysts (slow vs. fast). Genes in blue represent upregulation in slow blastocysts, genes in red represent upregulation in fast blastocysts. log2FoldCh Gene p-value p-Adj ange PDHB −1.425 0.000 0.000 MDH1 −1.206 0.000 0.000 APEX1 −1.193 0.000 0.000 OGDHL −3.417 0.000 0.002 PGK1 −0.942 0.000 0.002 GLS2 1.493 0.000 0.002 AICDA 1.171 0.001 0.005 ACO2 0.693 0.002 0.011 CS −0.660 0.002 0.011 SLC25A1 1.181 0.007 0.032 IDH3A −0.728 0.008 0.035 GSS 1.039 0.013 0.053 TET3 0.662 0.026 0.093 GLUD1 −0.450 0.032 0.108 SDHD −0.619 0.049 0.143 FH −0.547 0.054 0.149 OGDH 0.316 0.133 0.287 ACO1 −0.364 0.141 0.297 SDHC −0.335 0.149 0.311 LIG3 0.338 0.165 0.334 SUCLG −0.332 0.174 0.349 SDHA 0.297 0.210 0.396 SUCLA2 −0.324 0.248 0.439 DNMT1 0.266 0.279 0.486 IDH3B1 −0.269 0.296 0.503 SDHB −0.213 0.339 0.544 DNMT3B 0.181 0.386 0.598 APOBEC1 0.629 0.386 0.598 TDG 0.427 0.398 0.611 IDH3G 0.237 0.468 0.675 NEIL2 0.509 0.572 0.720 IDH2 0.298 0.571 0.720 DNMT3L 1.306 0.590 0.722 GLS 0.120 0.706 0.821 XRCC1 0.108 0.793 0.887 TET1 −0.028 0.879 0.919 DNMT3A 0.029 0.893 0.920 MBD4 −0.056 0.885 0.920 PDHX 0.033 0.890 0.920 SMUG1 0.053 0.936 0.954 TET2 −0.002 0.991 0.991 Table S2. -
Synthetic Lethality Between DNA Polymerase Epsilon and RTEL1 in Metazoan DNA Replication
Article Synthetic Lethality between DNA Polymerase Epsilon and RTEL1 in Metazoan DNA Replication Graphical Abstract Authors Roberto Bellelli, Jillian Youds, Valerie Borel, Jennifer Svendsen, Visnja Pavicic-Kaltenbrunner, Simon J. Boulton Correspondence [email protected] In Brief Bellelli et al. report that RTEL1 deficiency is synthetic lethal with the loss of pole-4 in C. elegans/hypomorphy of Pol epsilon. An analysis of replication dynamics in Rtel1À/À Pole4À/À mouse cells revealed a combination of dysfunctional fork progression and defective origin activation, which cooperatively drive incomplete genomic replication and genetic instability. Highlights d rtel-1 is synthetic lethal with the loss of DNA polymerase epsilon in C. elegans d rtel-1; pole-4 double mutants accumulate Rad51 and RPA foci and fail to replicate d Impaired DNA replication and genome instability in Rtel1 Pole4 knockout mouse cells d Rtel1 Pole4 double knockout mouse cells exhibit fork asymmetry and defective origin activation Bellelli et al., 2020, Cell Reports 31, 107675 May 26, 2020 ª 2020 The Author(s). https://doi.org/10.1016/j.celrep.2020.107675 ll ll OPEN ACCESS Article Synthetic Lethality between DNA Polymerase Epsilon and RTEL1 in Metazoan DNA Replication Roberto Bellelli,1,2,3 Jillian Youds,1,2 Valerie Borel,1 Jennifer Svendsen,1 Visnja Pavicic-Kaltenbrunner,1 and Simon J. Boulton1,4,* 1The Francis Crick Institute, 1 Midland Road, NW1 1AT London, UK 2These authors contributed equally 3Present address: Center for Cancer Cell and Molecular Biology, Barts Cancer Institute, Queen Mary University of London, Charterhouse Square, Barbican, EC1M 6BE London, UK 4Lead Contact *Correspondence: [email protected] https://doi.org/10.1016/j.celrep.2020.107675 SUMMARY Genome stability requires coordination of DNA replication origin activation and replication fork progression. -

Evidence Supporting Dissimilatory And
University of Massachusetts Amherst ScholarWorks@UMass Amherst Microbiology Department Faculty Publication Microbiology Series 2013 Evidence supporting dissimilatory and assimilatory lignin degradation in Enterobacter lignolyticus SCF1 Kristen DeAngelis University of Massachusetts Amherst, [email protected] Deepak Sharma University of Massachusetts Amherst Rebecca Varney University of Massachusetts Amherst Blake Simmons Joint BioEnergy Institute Nancy G. Isern Environmental Molecular Sciences Laboratory See next page for additional authors Follow this and additional works at: https://scholarworks.umass.edu/micro_faculty_pubs Part of the Microbiology Commons Recommended Citation DeAngelis, Kristen; Sharma, Deepak; Varney, Rebecca; Simmons, Blake; Isern, Nancy G.; Markillie, Lye Meng; Nicora, Carrie; Norbeck, Angela D.; Taylor, Ronald C.; Aldrich, Joshua T.; and Robinson, Errol W., "Evidence supporting dissimilatory and assimilatory lignin degradation in Enterobacter lignolyticus SCF1" (2013). Frontiers in Microbiology. 303. 10.3389/fmicb.2013.00280 This Article is brought to you for free and open access by the Microbiology at ScholarWorks@UMass Amherst. It has been accepted for inclusion in Microbiology Department Faculty Publication Series by an authorized administrator of ScholarWorks@UMass Amherst. For more information, please contact [email protected]. Authors Kristen DeAngelis, Deepak Sharma, Rebecca Varney, Blake Simmons, Nancy G. Isern, Lye Meng Markillie, Carrie Nicora, Angela D. Norbeck, Ronald C. Taylor, Joshua T. Aldrich, and Errol W. Robinson This article is available at ScholarWorks@UMass Amherst: https://scholarworks.umass.edu/micro_faculty_pubs/303 ORIGINAL RESEARCH ARTICLE published: 19 September 2013 doi: 10.3389/fmicb.2013.00280 Evidence supporting dissimilatory and assimilatory lignin degradation in Enterobacter lignolyticus SCF1 Kristen M. DeAngelis 1*, Deepak Sharma 1, Rebecca Varney 1, Blake Simmons 2,3, Nancy G. -

Kinase-Dead ATM Protein Is Highly Oncogenic and Can Be Preferentially Targeted by Topo
1 Kinase-dead ATM protein is highly oncogenic and can be preferentially targeted by Topo- 2 isomerase I inhibitors 3 4 Kenta Yamamoto1,2, Jiguang Wang3, Lisa Sprinzen1,2, Jun Xu5, Christopher J. Haddock6, Chen 5 Li1, Brian J. Lee1, Denis G. Loredan1, Wenxia Jiang1, Alessandro Vindigni6, Dong Wang5, Raul 6 Rabadan3 and Shan Zha1,4 7 8 1 Institute for Cancer Genetics, Department of Pathology and Cell Biology, College of Physicians 9 and Surgeons, Columbia University, New York City, NY 10032 10 2 Pathobiology and Molecular Medicine Graduate Program, Department of Pathology and Cell 11 Biology, Columbia University, New York City, NY 10032 12 3 Department of Biomedical Informatics and Department of Systems Biology, College of 13 Physicians & Surgeons, Columbia University, New York City, NY 10032 14 4 Division of Pediatric Oncology, Hematology and Stem Cell Transplantation, Department of 15 Pediatrics, College of Physicians & Surgeons, Columbia University, New York City, NY 10032 16 5 Skaggs School of Pharmacy & Pharmaceutical Sciences, University of California San Diego, 17 La Jolla, CA 92093 18 6 Edward A. Doisy Department of Biochemistry and Molecular Biology, Saint Louis University 19 School of Medicine, St. Louis, MO 63104 20 21 Short Title: Topo1 inhibitors target ATM mutated cancers 22 Key Words: ATM, missense mutations, Topo I inhibitors 23 24 Address Correspondence to: Shan Zha at [email protected] 25 26 1 27 ABSTRACT 28 Missense mutations in ATM kinase, a master regulator of DNA damage responses, are 29 found in many cancers, but their impact on ATM function and implications for cancer therapy are 30 largely unknown. -

POLD3 Is Haploinsufficient for DNA Replication in Mice
POLD3 is haploinsufficient for DNA replication in mice Matilde Murga1, Emilio Lecona1, Irene Kamileri2, Marcos Díaz3, Natalia Lugli2, Sotirios K. Sotiriou2, Marta E. Anton1, Juan Méndez3, Thanos D. Halazonetis2 and Oscar Fernandez-Capetillo1,4 1 Genomic Instability Group, Spanish National Cancer Research Centre, Madrid, Spain. 2Department of Molecular Biology, University of Geneva, Geneva, Switzerland. 3 DNA Replication Group, Spanish National Cancer Research Centre, Madrid, Spain. 4Science for Life Laboratories, Division of Translational Medicine and Chemical Biology, Department of Medical Biochemistry and Biophysics, Karolinska Institute, Stockholm, Sweden. Correspondence: O.F. ([email protected]) Contact: Oscar Fernandez-Capetillo Spanish National Cancer Research Centre (CNIO) Melchor Fernandez Almagro, 3 Madrid 28029, Spain Tel.: +34.91.732.8000 Ext: 3480 Fax: +34.91.732.8028 Email: [email protected] POLD3 deficient mice SUMMARY The Pold3 gene encodes a subunit of the Polδ DNA polymerase complex. Pold3 orthologues are not essential in Saccharomyces cerevisiae or chicken DT40 cells, but the Schizzosaccharomyces pombe orthologue is essential. POLD3 also has a specialized role in the repair of broken replication forks, suggesting that POLD3 activity could be particularly relevant for cancer cells enduring high levels of DNA replication stress. We report here that POLD3 is essential for mouse development and is also required for viability in adult animals. Strikingly, even Pold3+/- mice were born at sub-Mendelian ratios and, of those born, some presented hydrocephaly and had a reduced lifespan. In cells, POLD3 deficiency led to replication stress and cell death, which were aggravated by expression of activated oncogenes. Finally, we show that Pold3 deletion destabilizes all members of the Polδ complex, explaining its major role in DNA replication and the severe impact of its deficiency. -

Bioinformatics-Based Screening of Key Genes for Transformation of Liver
Jiang et al. J Transl Med (2020) 18:40 https://doi.org/10.1186/s12967-020-02229-8 Journal of Translational Medicine RESEARCH Open Access Bioinformatics-based screening of key genes for transformation of liver cirrhosis to hepatocellular carcinoma Chen Hao Jiang1,2, Xin Yuan1,2, Jiang Fen Li1,2, Yu Fang Xie1,2, An Zhi Zhang1,2, Xue Li Wang1,2, Lan Yang1,2, Chun Xia Liu1,2, Wei Hua Liang1,2, Li Juan Pang1,2, Hong Zou1,2, Xiao Bin Cui1,2, Xi Hua Shen1,2, Yan Qi1,2, Jin Fang Jiang1,2, Wen Yi Gu4, Feng Li1,2,3 and Jian Ming Hu1,2* Abstract Background: Hepatocellular carcinoma (HCC) is the most common type of liver tumour, and is closely related to liver cirrhosis. Previous studies have focussed on the pathogenesis of liver cirrhosis developing into HCC, but the molecular mechanism remains unclear. The aims of the present study were to identify key genes related to the transformation of cirrhosis into HCC, and explore the associated molecular mechanisms. Methods: GSE89377, GSE17548, GSE63898 and GSE54236 mRNA microarray datasets from Gene Expression Omni- bus (GEO) were analysed to obtain diferentially expressed genes (DEGs) between HCC and liver cirrhosis tissues, and network analysis of protein–protein interactions (PPIs) was carried out. String and Cytoscape were used to analyse modules and identify hub genes, Kaplan–Meier Plotter and Oncomine databases were used to explore relationships between hub genes and disease occurrence, development and prognosis of HCC, and the molecular mechanism of the main hub gene was probed using Kyoto Encyclopedia of Genes and Genomes(KEGG) pathway analysis.