How to Argue with a Racist: History, Science, Race and Reality
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

BOSNIA and HERZEGOVINA MINISTRY of FOREIGN AFFAIRS Department of Diplomatic Protocol
BOSNIA AND HERZEGOVINA MINISTRY OF FOREIGN AFFAIRS Department of Diplomatic Protocol DIPLOMATIC AND CONSULAR CORPS AND INTERNATIONAL ORGANIZATIONS IN BOSNIA AND HERZEGOVINA April 2019 1 2 C O N T E N T S Order of Precedence among the Heads of Diplomatic Missions and dates of presentation of credentials 9 Heads of Diplomatic Representation Offices 11 Diplomatic Missions accredited to Bosnia and Herzegovina * - Non Resident ALBANIA* 15 ALGERIA * 17 ANGOLA * 19 ARGENTINA * 21 ARMENIA* 23 AUSTRALIA * 25 AUSTRIA 27 AZERBAIJAN* 29 BAHRAIN* 31 BANGLADESH * 33 BELARUS * 35 BELGIUM * 37 BRAZIL 39 BRUNEI DARUSSALAM* 41 BULGARIA 43 BURKINA FASO * 45 CANADA* 47 CHILE * 49 CHINA 51 COSTA RICA* 53 CROATIA 55 CUBA * 57 CYPRUS* 59 CZECH REPUBLIC 61 DENMARK* 63 ECUADOR* 65 3 EGYPT 67 ESTONIA * 69 ETHIOPIA* 71 FINLAND * 73 FRANCE 75 GEORGIA* 77 GERMANY 79 GHANA* 83 GREECE 85 HOLY SEE 87 HUNGARY 89 ICELAND * 91 INDIA * 93 INDONESIA 95 IRAN 97 IRAQ* 99 IRELAND * 101 ISRAEL * 103 ITALY 105 JAPAN 107 JORDAN * 109 KAZAKHSTAN* 111 KOREA (Democratic People's Republic) * 113 KOREA (Republic of Korea) * 115 KUWAIT 117 LATVIA * 119 LITHUANIA* 121 LIBYA 123 LUXEMBOURG* 125 MALAYSIA 127 MALI * 129 MALTA * 131 MAURITANIA* 133 MEXICO * 135 4 MOLDOVA * 137 MONGOLIA* 139 MONTENEGRO 141 MOROCCO * 143 NETHERLANDS 145 NEW ZEALAND * 147 NIGERIA * 149 NORTH MACEDONIA 151 NORWAY 153 OMAN * 155 PAKISTAN 157 PALESTINE 159 PERU * 161 PHILIPPINES * 163 POLAND 165 PORTUGAL* 167 QATAR 169 ROMANIA 171 RUSSIA 173 SAN MARINO 177 SAUDI ARABIA 179 SERBIA 183 SYRIA * 185 SLOVAKIA 187 SLOVENIA -

2019 Tokyo Marathon Statistical Information
2019 Tokyo Marathon Statistical Information Tokyo Marathon All Time list Performance Time Performers Name Nat Place Date 1 2:03:58 1 Wilson Kipsang KEN 1 26 Feb 2017 2 2:05:30 2 Dickson Chumba KEN 1 25 Feb 2018 3 2:05:42 Dickson Chumba 1 23 Feb 2014 4 2:05:51 3 Gideon Kipketer KEN 2 26 Feb 2017 5 2:05:57 4 Tadese Tola ETH 2 23 Feb 2014 6 2:06:00 5 Endeshaw Negesse ETH 1 22 Feb 2015 7 2:06:11 6 Yuta Shitara JPN 2 25 Feb 2018 8 2:06:25 Dickson Chumba 3 26 Feb 2017 9 2:06:30 7 Sammy Kitwara KEN 3 23 Feb 2014 10 2:06:33 8 Stephen Kiprotich UGA 2 22 Feb 2015 11 2:06:33 9 Amos Kipruto KEN 3 25 Feb 2018 12 2:06:34 Dickson Chumba 3 22 Feb 2015 13 2:06:42 10 Evans Chebet KEN 4 26 Feb 2017 14 2:06:47 Gideon Kipketer 4 25 Feb 2018 15 2:06:50 11 Dennis Kimetto KEN 1 24 Feb 2013 16 2:06:54 12 Hiroto Inoue JPN 5 25 Feb 2018 17 2:06:56 13 Feyisa Lilesa ETH 1 28 Feb 2016 18 2:06:58 14 Michael Kipyego KEN 2 24 Feb 2013 19 2:06:58 Michael Kipyego 4 23 Feb 2014 20 2:07:05 15 Peter Some KEN 5 23 Feb 2014 21 2:07:20 16 Shumi Dechasa BRN 4 22 Feb 2015 22 2:07:22 Peter Some 5 22 Feb 2015 23 2:07:23 17 Viktor Röthlin SUI 1 17 Feb 2008 24 2:07:25 18 Markos Geneti ETH 6 22 Feb 2015 25 2:07:30 Feyisa Lilesa 6 25 Feb 2018 26 2:07:33 19 Bernard Kipyego KEN 2 28 Feb 2016 27 2:07:34 Dickson Chumba 3 28 Feb 2016 28 2:07:35 20 Hailu Mekonnen ETH 1 27 Feb 2011 29 2:07:37 Michael Kipyego 1 26 Feb 2012 30 2:07:37 21 Geoffrey Kamworor Kipsang KEN 6 23 Feb 2014 31 2:07:39 22 Masato Imai JPN 7 22 Feb 2015 32 2:07:39 23 Alfers Lagat KEN 5 26 Feb 2017 33 2:07:40 24 Deresa Chimsa -

Baku 2015 European Games – Men Athlete Profiles 1 49KG
Baku 2015 European Games – Men Athlete Profiles 49KG – ARTYOM ALEKSANYAN – ARMENIA (ARM) Date Of Birth : 24/06/1996 Place Of Birth : Martuni Height : 159cm Coach : Fedya Aleksanyan Club : YSSCOR Residence : Ararat Stance : Orthodox Number of training hours : 20 in a week Number of bouts : 54 Began boxing : 2006 2015 – Liventsev Memorial Tournament (Minsk, BLR) 7th place – 49KG Lost to Abdulla Kuchkarov (UZB) 3:0 in the quarter-final 2015 – Armenian National Championships 2nd place – 49KG Lost to Artur Hovhannisyan (ARM) 2:1 in the final; Won against Harutyun Harutyunyan (ARM) 3:0 in the semi-final 2014 – Kutaisi International Youth Tournament (Kutaisi, GEO) 1st place – 49KG 2014 – Armenian Youth National Championships 3rd place – 49KG Lost to Artyom Hovhannisyan (ARM) 2:1 in the semi-final 49KG – SALMAN ALIZADA – AZERBAIJAN (AZE) Date Of Birth : 01/12/1993 Place Of Birth : Baku Height : 163cm Coach : Nazim Guseynov Club : Gilan Residence : Baku Hobbies : Travelling; Photography Began boxing : 2004 2015 – Great Silk Way Tournament (Baku, AZE) 6th place – 49KG Lost to Tinko Banabakov (BUL) 2:1 in the quarter-final 2015 – Bocskai Memorial Tournament (Debrecen, HUN) 2nd place – 52KG Lost to Masud Yusifzada (AZE) 2:0 in the final; Won against Nurlan Askhar (MGL) 2:1 in the semi-final; Won against Kuanysh Sabyr (KAZ) 3:0 in the quarter-final 2014 – World Cup of Petroleum Countries (Beloyarskiy, RUS) 1st place – 49KG Won against Abdulla Kuchkarov (UZB) 3:0 in the final; Won against Turat Osmonov (KGZ) 2:0 in the semi-final; Won against Andrei -

Biology Things to Do
Post 16 Biology things to do: Competition: The Royal Society of Biology has a photography competition with the theme ‘our changing world’ more information can be found by following the link. https://www.rsb.org.uk/get-involved/rsb-competitions/photography-competition The prize for the winner of the under 18 competition is £500 Reading: We have split these reading lists into 3 categories, this is to give you an idea of how ‘easy’ the reading is – for example Bill Bryson’s ‘The Body’ whilst very factual and interesting is designed for the general population to read. On the other hand ‘Evolution in 4 dimensions’ is designed with a biologist in mind – if evolution is your thing – give it a go! Category 1 are the easiest reads and category 3 are the most difficult – but all worthwhile. Category 1 Category 2 Category 3 ‘The Body’ https://thebiologist.rsb.org.uk/biologist ‘Evolution in 4 dimensions’ Bill Bryson Eva Jablonka & Marion J.Lamb ‘A short history of nearly ‘Darwin comes to Town’ ‘The Epigenetics everything’ Memo Schilthuizen Revolution’ Bill Bryson Nessa Carey ‘Sapiens’ ‘The immortal life of Henrietta Lacks’ ‘The Cell: Discovering the Yuval Noah Harari Rebecca Skloot microscopic World that Determines our Health, Consciousness, and Our Future’ Joshua Z.Rappoport PhD, Barry Abrams et al. ‘Homo Deus’ ‘Evolution: The Human Story, 2nd ‘Life: an unauthorized Yuval Noah Harari edition’ biography’ Alice Roberts Richard Fortey ‘The Selfish Gene’ ‘The soul of an octopus’ Richard Dawkins Sy Montgomery ‘The Ancestor’s Tale’ ‘The Gene: An intimate history’ Richard Dawkins and Yan Siddhartha Mukherjee Wong ‘This is going to hurt’ ‘In the shadow of man’ Adam Kay Jane Goodall ‘Human Universe’ ‘The Book of Humans’ Brian Cox Adam Rutherford ‘Life on Earth’ David Attenborough ‘Anatomy and physiology for dummies’ Tasks Option 1: https://pbiol.rsb.org.uk/cells-to-systems/support-and-movement/observing-earthworm-locomotion The idea is to find Earthworms and compare their movement on different surfaces. -

AIBA Youth World Boxing Championships Yerevan 2012 Athletes Biographies
AIBA Youth World Boxing Championships Yerevan 2012 Athletes Biographies 49KG – HAYRIK NAZARYAN – ARMENIA (ARM) Date Of Birth : 30/08/1995 Club : Working Shift Sport Company Coach : Marat Karoyan Residence : Yerevan Number of bouts : 60 Began boxing : 2002 2012 – Klichko Brothers Youth Tournament (Berdichev, UKR) 6th place – 49KG Lost to Sultan Abduraimov (KAZ) 12:3 in the quarter-final; Won against Danilo Pleshkov (UKR) AB 2nd round in the first preliminary round 2012 – Armenian Youth National Championships 1st place – 49KG Won against Andranik Peleshyan (ARM) by points in the final; Won against Taron Petrosyan (ARM) by points in the semi-final 2012 – Pavlyukov Youth Memorial Tournament (Anapa, RUS) 7th place – 49KG Lost to Keith Flavin (IRL) 30:6 in the quarter-final 2011 – AIBA Junior World Championships (Astana, KAZ) 7th place – 46KG Lost to Georgian Tudor (ROM) 15:14 in the quarter-final; Won against Dmitriy Asanov (BLR) 22:14 in the first preliminary round 2011 – European Junior Championships (Keszthely, HUN) 5th place – 46KG Lost to Timur Pirdamov (RUS) 17:4 in the quarter-final; Won against Zsolt Csonka (HUN) RSC 2nd round in the first preliminary round 2011 – Armenian Junior National Championships 1st place – 46KG 49KG – ROBERT TRIGG – AUSTRALIA (AUS) Date Of Birth : 03/01/1994 Place Of Birth : Mount Gambier Height : 154cm Club : Mt. Gambier Boxing Club Coach : Colin Cassidy Region : South Australia Began boxing : 2010 2012 – Oceanian Youth Championships (Papeete, TAH) 1st place – 49KG Won against Martin Dexon (NRU) by points -

CCBE-INFO May 2007 / N°18
CCBE-INFO May 2007 / N°18 Représentant les avocats d’Europe Representing Europe’s lawyers Content Services Directive . .2 Money Laundering . .2 Commission report . .2 Competition . .2 Conveyancing services study . .2 CCBE Human Rights Award for European Lawyers . .3 What is the CCBE Human Rights Award? . .3 Procedure for the nomination of candidates for the CCBE Human Rights Award . .3 Training . .4 PECO . .4 CCBE Membership – Romania – Legal Advisers . .4 Ukraine . .4 Moldova . .4 Access to Justice Committee: The attachment of bank accounts . .4 Criminal law . .5 CCBE letter on procedural safeguards . .5 Provision and quality of legal interpreting and translation . .5 Corporate Social responsibility . .6 Questionnaire on CSR . .6 European Contract Law . .6 Family Law . .7 GATS . .7 Mutual Recognition Agreement (MRA) with the American Bar Association . .7 International Private Law . .7 IT-Law . .8 Establishment of EU lawyers in Luxembourg - Update . .8 Multi-jurisdictional Law Firms Committee . .8 Professional Indemnity (PI) Insurance . .9 Social Security . .9 Editor: Peter Mc Namee Editorial Board: Birgit Beger Sieglinde Gamsjäger Karine Métayer Peter Mc Namee Editorial Secretaries: Maxime Servotte (translator) Sophie Testaert (layout) ccbe-info published 4 times / year Subscription is free of charge by send- ing an e-mail to: [email protected] 1 ccbe-info n° 18/2007 Services Directive In order to help Bars and Law Societies with the implementation of the Directive on Services in the Internal Market (O.J. L 376/36 du 27.12.2006 p. 36), the CCBE has adopted an internal commentary on the Directive. The commentary, which is not public at this stage, is meant to be an evolving document, which might be further completed and amended. -

Radio 4 Listings for 29 February – 6 March 2020 Page 1 of 14
Radio 4 Listings for 29 February – 6 March 2020 Page 1 of 14 SATURDAY 29 FEBRUARY 2020 Series 41 SAT 10:30 The Patch (m000fwj9) Torry, Aberdeen SAT 00:00 Midnight News (m000fq5n) The Wilberforce Way with Inderjit Bhogal National and international news from BBC Radio 4 The random postcode takes us to an extraordinary pet shop Clare Balding walks with Sikh-turned-Methodist, Inderjit where something terrible has been happening to customers. Bhogal, along part of the Wilberforce Way in East Yorkshire. SAT 00:30 The Crying Book, by Heather Christle Inderjit created this long distance walking route to honour Torry is a deprived area of Aberdeen, known for addiction (m000fq5q) Wilberforce who led the campaign against the slave trade. They issues. It's also full of dog owners. In the local pet shop we Episode 5 start at Pocklington School, where Wilberforce studied, and discover Anna who says that a number of her customers have ramble canal-side to Melbourne Ings. Inderjit Bhogal has an died recently from a fake prescription drug. We wait for her Shedding tears is a universal human experience, but why and extraordinary personal story: Born in Kenya he and his family most regular customer, Stuart, to help us get to the bottom of it how do we cry? fled, via Tanzania, to Dudley in the West Midlands in the early - but where is he? 1960s. He couldn’t find anywhere to practice his Sikh faith so American poet Heather Christle has lost a dear friend to suicide started attending his local Methodist chapel where he became Producer/presenter: Polly Weston and must now reckon with her own depression. -

Even Scientists Can't Predict the Future – Or Can They? Edited by Jim Al
EVEN SCIEntists can’t prEDICT THE FUTURE – OR CAN THEY? Edited by Jim Al-Khalili PROFILE BOOKS What's Next?.indd 3 30/08/2017 16:25 First published in Great Britain in 2017 by Profile Books Ltd 3 Holford Yard Bevin Way London wc1x 9hd www.profilebooks.com 10 9 8 7 6 5 4 3 2 1 Selection, introduction and Chapter 18 (‘Teleportation and Time Travel’) copyright © Jim Al-Khalili 2017 Other chapters copyright of the author in each case © Philip Ball, Margaret A. Boden, Naomi Climer, Lewis Dartnell, Jeff Hardy, Winfried K. Hensinger, Adam Kucharski, John Miles, Anna Ploszajski, Aarathi Prasad, Louisa Preston, Adam Rutherford, Noel Sharkey, Julia Slingo, Gaia Vince, Mark Walker, Alan Woodward 2017 The moral right of the authors has been asserted. All rights reserved. Without limiting the rights under copyright reserved above, no part of this publication may be reproduced, stored or introduced into a retrieval system, or transmitted, in any form or by any means (electronic, mechanical, photocopying, recording or otherwise), without the prior written permission of both the copyright owner and the publisher of this book. All reasonable efforts have been made to obtain copyright permissions where required. Any omissions and errors of attribution are unintentional and will, if notified in writing to the publisher, be corrected in future printings. A CIP catalogue record for this book is available from the British Library. ISBN 978 1 78125 895 8 eISBN 978 1 78283 376 5 Text design by Sue Lamble Typeset in Sabon by MacGuru Ltd Printed and bound in Britain -

Applied Science (Equivalent to One a Level)
Applied Science (equivalent to one A Level) This is a programme of activities and resources to prepare you to start BTEC Applied Science in September. It is aimed to be used throughout the remainder of the Summer term and over the Summer Holidays to ensure you are ready to start your course in September. Book Recommendations The books below are all popular science books and great for extending your understanding of Science. Bryson and Goldacre are available as ebooks from Hampshire library service. https://www.hants.gov.uk/librariesandarchives/library/whatyoucanborrow/ebooksaudi obooks A Short History of Nearly Everything A whistle-stop tour through many aspects of history from the Big Bang to now. This is a really accessible read that will re-familiarise you with common concepts and introduce you to some of the more colourful characters from the history of science! Available at amazon.co.uk Periodic Tales: The Curious Lives of the Elements (Paperback) Hugh AlderseyWilliams ISBN-10: 0141041455 http://bit.ly/pixlchembook1 This book covers the chemical elements, where they come from and how they are used. There are loads of fascinating insights into uses for chemicals you would have never even thought about. Bad Science (Paperback) Ben Goldacre ISBN-10: 000728487X http://bit.ly/pixlchembook3 Here Ben Goldacre takes apart anyone who published bad / misleading or dodgy science – this book will make you think about everything the advertising industry tries to sell you by making it sound ‘sciency’. TV and film Recommendations You won’t find Jurassic Park on this list, but they are all great watching for a rainy day. -

Dangerous Teen Fads II
3/12/2015 Dangerous Teen Fads II Marilyn GrechusGrechus,, Ph.D. University of Central Missouri [email protected] Children and teens are engaging in increasingly risky behaviors. It is important for us, as educators, to understand and stay up-to-date on these dangerous teen fads. Remember: Knowledge is Power. 1 3/12/2015 Dangerous Teen Fads II This presentation is a followfollow----upup to the Dangerous Teen Fads presentation from last year’s national convention. I’ve included a review of many of last year’s fads for your information. This Review is presented in an abbreviated fashion. The represents some of the consequences caused by the behavior. To locate more info on any of the trends presented, you can search youtube videos by name of the behavior or search Dangerous Teen Trends. 2 3/12/2015 Review Vodka Eyeballing Pouring vodka into one’s eye Burning, blindness Vodka Tampons Inserted into vagina or anus Alcohol poisoning, bleeding Review Neknominate Online drinking game, dare friend to top it Do stupid, dangerous things Pharm Parties Prescription drugs in a bowl, take handful Death RoboRobo----TrippingTripping Ingesting large amounts of dextromethorphan Hallucinations, confusion, alcohol intoxification 3 3/12/2015 Review Inhalant Abuse Huffing, Sniffing, Bagging Suffocation, seizures, heart failure, death Bath Salts Designer drug Paranoia, hallucinations, Violence, Suicide Biting New ‘hickey’, Pain is a rush Hepatitis, syphilis, HIV/AIDS, infection Review Online Pictures / Sexting Posting or sending sexually explicit pictures Bad -

Hell on Earth
01 CHAPTER 3 02 03 04 Hell on Earth 05 06 07 08 09 10 11 12 “Long is the way, and hard, that out of hell leads up to 13 light.” 14 John Milton, Paradise Lost 15 16 17 f you want to construct a picture of the earth on which life first 18 emerged, think about how we’ve named it. There are four geo- 19 logical eons spanning the earth’s 4, 540-million-year existence. 20 IThe most recent three names reflect our planet’s propensity for liv- 21 ing things, all referring to stages of life. The second eon is called the 22 Archean, which rather confusingly translates as “origins.” The third 23 eon is the Proterozoic, roughly translating from Greek as “earlier 24 life”; the current eon, the Phanerozoic, started around 542 million 25 years ago and means “visible life.” 26 But the first eon, the period from the formation of the earth up 27 to 3. 8 billion years ago, is called the Hadean, derived from Hades, 28 the ancient Greek version of hell. 29 Life does not merely inhabit this planet; it has shaped it and is 30 part of it. Not just in the current era of man-made climate change, S31 but all through life’s history on Earth, life has affected the rocks N32 9781617230059_Creation_TX_p1-278.indd 61 26/02/13 7:51 PM 62 CREATION 01 below our feet and the sky above us. And necessarily, the origin of 02 life is inseparable from the fury of the formation of the earth in the 03 first place. -
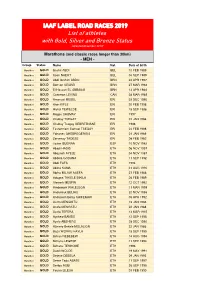
UPDATED List of Athletes with Label Status 2019
IAAF LABEL ROAD RACES 2019 List of athletes with Gold, Silver and Bronze Status (Updated November 2019) Marathons (and classic races longer than 30km) - MEN - Group Status Name Nat. Date of birth Marathon GOLD Bashir ABDI BEL 10 FEB 1989 Marathon GOLD Koen NAERT BEL 03 SEP 1989 Marathon GOLD Abdi Ibrahim ABDO BRN 24 APR 1997 Marathon GOLD Benson SEUREI BRN 27 MAR 1984 Marathon GOLD El Hassan EL ABBASSI BRN 13 APR 1984 Marathon GOLD Cameron LEVINS CAN 28 MAR 1989 Marathon GOLD Amanuel MESEL ERI 29 DEC 1990 Marathon GOLD Aron KIFLE ERI 20 FEB 1998 Marathon GOLD Hiskel TEWELDE ERI 15 SEP 1986 Marathon GOLD Mogos SHUMAY ERI 1997 Marathon GOLD Okubay TSEGAY ERI 01 JAN 1986 Marathon GOLD Okubay Tsegay GEBRETNSAE ERI 1986 Marathon GOLD Tesfamriam Samuel TSEGAY ERI 24 FEB 1988 Marathon GOLD Yohanes GHEBREGERGIS ERI 01 JAN 1989 Marathon GOLD Zersenay TADESE ERI 08 FEB 1982 Marathon GOLD Javier GUERRA ESP 10 NOV 1983 Marathon GOLD Abadi HADIS ETH 06 NOV 1997 Marathon GOLD Abayneh AYELE ETH 04 NOV 1987 Marathon GOLD Abdela GODANA ETH 11 SEP 1992 Marathon GOLD Abdi FUFA ETH 1994 Marathon GOLD Abera KUMA ETH 31 AUG 1990 Marathon GOLD Abrha MILAW ASEFA ETH 27 FEB 1988 Marathon GOLD Adugna TAKELE BIKILA ETH 26 FEB 1989 Marathon GOLD Afewerk MESFIN ETH 12 OCT 1992 Marathon GOLD Amdework WALELEGN ETH 11 MAR 1999 Marathon GOLD Andamlak BELIHU ETH 20 NOV 1998 Marathon GOLD Andualem Belay SHIFERAW ETH 05 APR 1992 Marathon GOLD Asefa MENGISTU ETH 18 JAN 1985 Marathon GOLD Asefa MENGSTU ETH 22 JAN 1988 Marathon GOLD Asefa TEFERA ETH 14 MAR 1997 Marathon GOLD