Experimental Models for Fungal Keratitis: an Overview of Principles and Protocols
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
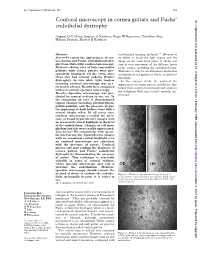
Confocal Microscopy in Cornea Guttata and Fuchs' Endothelial Dystrophy
Br J Ophthalmol 1999;83:185–189 185 Confocal microscopy in cornea guttata and Fuchs’ Br J Ophthalmol: first published as 10.1136/bjo.83.2.185 on 1 February 1999. Downloaded from endothelial dystrophy Auguste G-Y Chiou, Stephen C Kaufman, Roger W Beuerman, Toshihiko Ohta, Hisham Soliman, Herbert E Kaufman Abstract conventional imaging methods.3–13 Because of Aims—To report the appearances of cor- its ability to focus the light source and the nea guttata and Fuchs’ endothelial dystro- image on the same focal plane, it allows real phy from white light confocal microscopy. time in vivo assessment of the diVerent layers Methods—Seven eyes of four consecutive of the cornea, including the endothelial layer. patients with cornea guttata were pro- Therefore, it may be an alternative method in spectively examined. Of the seven eyes, evaluating cornea guttata or Fuchs’ endothelial three also had corneal oedema (Fuchs’ dystrophy. dystrophy). In vivo white light tandem In the current study, we analysed the scanning confocal microscopy was per- appearances of cornea guttata and Fuchs’ dys- formed in all eyes. Results were compared trophy from confocal microscopy and compare with non-contact specular microscopy. the technique with non-contact specular mi- Results—Specular microscopy was pre- croscopy. cluded by corneal oedema in one eye. In the remaining six eyes, it demonstrated typical changes including pleomorphism, polymegathism, and the presence of gut- tae appearing as dark bodies, some with a central bright reflex. In all seven eyes, confocal microscopy revealed the pres- ence of round hyporeflective images with an occasional central highlight at the level of the endothelium. -

Curvularia Keratitis*
09 Wilhelmus Final 11/9/01 11:17 AM Page 111 CURVULARIA KERATITIS* BY Kirk R. Wilhelmus, MD, MPH, AND Dan B. Jones, MD ABSTRACT Purpose: To determine the risk factors and clinical signs of Curvularia keratitis and to evaluate the management and out- come of this corneal phæohyphomycosis. Methods: We reviewed clinical and laboratory records from 1970 to 1999 to identify patients treated at our institution for culture-proven Curvularia keratitis. Descriptive statistics and regression models were used to identify variables associ- ated with the length of antifungal therapy and with visual outcome. In vitro susceptibilities were compared to the clini- cal results obtained with topical natamycin. Results: During the 30-year period, our laboratory isolated and identified Curvularia from 43 patients with keratitis, of whom 32 individuals were treated and followed up at our institute and whose data were analyzed. Trauma, usually with plants or dirt, was the risk factor in one half; and 69% occurred during the hot, humid summer months along the US Gulf Coast. Presenting signs varied from superficial, feathery infiltrates of the central cornea to suppurative ulceration of the peripheral cornea. A hypopyon was unusual, occurring in only 4 (12%) of the eyes but indicated a significantly (P = .01) increased risk of subsequent complications. The sensitivity of stained smears of corneal scrapings was 78%. Curvularia could be detected by a panfungal polymerase chain reaction. Fungi were detected on blood or chocolate agar at or before the time that growth occurred on Sabouraud agar or in brain-heart infusion in 83% of cases, although colonies appeared only on the fungal media from the remaining 4 sets of specimens. -

Original Article
Clinical and Experimental Ophthalmology 2007; 35: 124–130 doi:10.1111/j.1442-9071.2006.01405.x Original Article Fungal keratitis in Melbourne Prashant Bhartiya FRCS,1,2 Mark Daniell FRANZCO,1,2 Marios Constantinou BScHons BOrth,1,2 FM Amirul Islam PhD1,2 and Hugh R Taylor AC FRANZCO1,2 1Centre for Eye Research Australia, University of Melbourne, and 2Corneal Clinic, Royal Victorian Eye and Ear Hospital, Melbourne, Victoria, Australia ABSTRACT INTRODUCTION Background: Description of the clinical and microbiolog- Fungal keratitis is a potentially blinding ocular disease. The ical spectrum of fungal keratitis at a tertiary eye care hos- incidence of fungal keratitis varies widely throughout the pital in Melbourne, Australia. world. A report from India showed that nearly 50% of all corneal ulcers were caused by fungi.1 This high prevalence Methods: Retrospective review of all patients with keratitis of fungal pathogens in south India is significantly greater with positive fungal cultures from corneal or associated than that found in similar studies in Nepal (17%),2 samples presenting to the Royal Victorian Eye and Ear Hos- Bangladesh (36%)3 and south Florida (35%).4 Several large pital, Melbourne, Australia from July 1996 to May 2004. studies on fungal keratitis have been published from North 4–12 Demographic data, predisposing factors, features on pre- and South America, Africa and the Indian subcontinent. However, there is a paucity of data on the spectrum of fungal sentation, management, outcomes and microbiological data keratitis in patients from Australia. This study reviewed a were collected and analysed. series of patients with keratitis who had fungal growth on Results: The study included 56 eyes of 56 patients. -

Two Cases of Keratomycosis Caused by Fusarium Solani: Therapeutic Management Fili S*, Schilde T, Perdikakis G and Kohlhaas M
Case Report iMedPub Journals Journal of Eye & Cataract Surgery 2016 http://www.imedpub.com/ Vol.2 No.1:9 ISSN 2471-8300 DOI: 10.21767/2471-8300.100009 Two Cases of Keratomycosis Caused by Fusarium Solani: Therapeutic Management Fili S*, Schilde T, Perdikakis G and Kohlhaas M Department of Ophthalmology, St.-Johannes-Hospital, Johannesstrasse, Dortmund, Germany *Corresponding author: Fili S, Department of Ophthalmology, St.-Johannes-Hospital, Johannesstrasse, Dortmund, Germany, Tel: 004915206428718; E-mail: [email protected] Received date: December 25, 2015; Accepted date: March 10, 2016; Published date: March 14, 2016 Citation: Fili S, Schilde T, Perdikakis G, Kohlhaas M (2015) Welcome Letter. J Eye Cataract Surg 2:9. doi: 10.21767/2471-8300.100009 Copyright: © 2016 Fili S, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited. Abbreviations Abstract EUCAST: European Committee on Antimicrobial susceptibility Testing, MIC: Minimum inhibitory concentration Backgrounds: To report 2 cases of multidrug-resistant fungal keratitis caused by Fusarium solani. The patients underwent a different conservative and surgical therapy Introduction including in both of them a keratoplasty à chaud. Keratomycosis is the greek word equivalent to fungal keratitis, Methods and Findings: A 21-years-old male and a 26-years- and it refers to a corneal infection caused by fungi which can old female patient, who both used soft contact lenses end up in severe loss of visual acuity or even enucleation. Fungal developed corneal infiltrates. In the first case the diagnosis keratitis is encountered in tropical climates and in populations was delayed because multiple microbiological examinations related to land cultivation. -

Review the Global Incidence and Diagnosis of Fungal Keratitis
Review The global incidence and diagnosis of fungal keratitis Lottie Brown, Astrid K Leck, Michael Gichangi, Matthew J Burton, David W Denning Fungal keratitis is a severe corneal infection that often results in blindness and eye loss. The disease is most prevalent Lancet Infect Dis 2020 in tropical and subtropical climates, and infected individuals are frequently young agricultural workers of low Published Online socioeconomic status. Early diagnosis and treatment can preserve vision. Here, we discuss the fungal keratitis October 22, 2020 diagnostic literature and estimate the global burden through a complete systematic literature review from January, 1946 https://doi.org/10.1016/ S1473-3099(20)30448-5 to July, 2019. An adapted GRADE score was used to evaluate incidence papers—116 studies provided the incidence of University of Manchester, fungal keratitis as a proportion of microbial keratitis and 18 provided the incidence in a defined population. We Manchester, UK (L Brown MSc, calculated a minimum annual incidence estimate of 1 051 787 cases (736 251–1 367 323), with the highest rates in Asia Prof D W Denning FRCP); and Africa. If all culture-negative cases are assumed to be fungal, the annual incidence would be 1 480 916 cases International Centre for Eye (1 036 641–1 925 191). In three case series, 8–11% of patients had to have the eye removed, which represents an annual Health, London School of Hygiene & Tropical Medicine, loss of 84 143–115 697 eyes. As fungal keratitis probably affects over a million people annually, an inexpensive, simple London, UK (A K Leck PhD, diagnostic method and affordable treatment are needed in every country. -

Topical Corticosteroids and Fungal Keratitis: a Review of the Literature and Case Series
Journal of Clinical Medicine Review Topical Corticosteroids and Fungal Keratitis: A Review of the Literature and Case Series Karl Anders Knutsson 1,*, Alfonso Iovieno 2,3, Stanislav Matuska 1, Luigi Fontana 2 and Paolo Rama 1 1 Cornea and Ocular Surface Unit, San Raffaele Scientific Institute, 20132 Milan, Italy; [email protected] (S.M.); [email protected] (P.R.) 2 Arcispedale Santa Maria Nuova—IRCCS, 42123 Reggio Emilia, Italy; [email protected] (A.I.); [email protected] (L.F.) 3 Department of Ophthalmology, University of British Columbia, Vancouver, BC V6T 1Z, Canada * Correspondence: [email protected] or [email protected]; Tel./Fax: +39-022-6432-648 Abstract: The management of fungal keratitis is complex since signs and symptoms are subtle and ocular inflammation is minimal in the preliminary stages of infection. Initial misdiagnosis of the condition and consequent management of inflammation with corticosteroids is a frequent occurrence. Topical steroid use is considered to be a principal factor for development of fungal keratitis. In this review, we assess the studies that have reported outcomes of fungal keratitis in patients receiving steroids prior to diagnosis. We also assess the possible rebound effect present when steroids are abruptly discontinued and the clinical characteristics of three patients in this particular clinical scenario. Previous reports and the three clinical descriptions presented suggest that in fungal keratitis, discontinuing topical steroids can induce worsening of clinical signs. In these cases, we recommend to slowly taper steroids and continue or commence appropriate antifungal therapy. Citation: Knutsson, K.A.; Iovieno, A.; Keywords: fungal keratitis; topical corticosteroids; topical steroids; rebound effect Matuska, S.; Fontana, L.; Rama, P. -

Fusarium Keratitis and Corneal Collagen Cross
FUSARIUM KERATITIS AND SURGERY REFRACTIVE CORNEAL COLLAGEN CROSS-LINKING BY MINAS CORONEO, AO, BSC(MED), MBBS, MSC SYD, MD, MS, UNSW, FRACS, FRANZCO; RAJESH FOGLA, DNB, FRCS(EDIN), MMED(OPHTH); WILLIAM B. TRATTLER, MD; ASHIYANA NARIANI, MD, MPH; COMPLEX CASE MANAGEMENT COMPLEX GARGI KHARE VORA, MD; AND ALAN N. CARLSON, MD CASE PRESENTATION A 42-year-old white man is referred to the Duke University Eye Center Cornea Service for a central corneal ulcer with a hypopyon in his right eye. The patient sustained the ocular injury while mowing the lawn, with debris getting into the eye while he was wearing contact lenses. He was diagnosed with culture-positive Fusarium species by the referring ophthalmologist and was treat- ed with oral voriconazole 200 mg twice daily and frequent topical natamycin 5% and voriconazole 10 mg/mL. The patient under- went epithelium-off corneal collagen cross-linking (CXL) approxi- Figure 1. Initial evaluation of the eye with a Fusarium corneal mately 4 weeks after diagnosis of the ulcer and was treated with infiltrate and hypopyon. a loteprednol steroid taper after the procedure. His condition subsequently progressed, with increasing eye pain, a nonhealing epithelial defect, and a worsening corneal infiltrate. Upon evaluation, the patient has a large corneal infiltrate with necrotic stroma, which is approaching the limbus, and a hypopyon (Figure 1). His UCVA measures 20/70-1. B-scan ultrasound of the right eye shows no evidence of posterior segment involvement. Reculturing of the corneal infiltrate is negative for bacteria, fungus, and Acanthamoeba. Confocal microscopy reveals no evidence of hyphae or cysts. -

Herpetic Corneal Infections
FocalPoints Clinical Modules for Ophthalmologists VOLUME XXVI, NUMBER 8 SEPTEMBER 2008 (MODULE 2 OF 3) Herpetic Corneal Infections Sonal S. Tuli, MD Reviewers and Contributing Editors Consultants George A. Stern, MD, Editor for Cornea & External Disease James Chodosh, MD, MPH Kristin M. Hammersmith, MD, Basic and Clinical Science Course Faculty, Section 8 Kirk R. Wilhelmus, MD, PhD Christie Morse, MD, Practicing Ophthalmologists Advisory Committee for Education Focal Points Editorial Review Board George A. Stern, MD, Missoula, MT Claiming CME Credit Editor in Chief, Cornea & External Disease Thomas L. Beardsley, MD, Asheville, NC Academy members: To claim Focal Points CME cred- Cataract its, visit the Academy web site and access CME Central (http://www.aao.org/education/cme) to view and print William S. Clifford, MD, Garden City, KS Glaucoma Surgery; Liaison for Practicing Ophthalmologists Advisory your Academy transcript and report CME credit you Committee for Education have earned. You can claim up to two AMA PRA Cate- gory 1 Credits™ per module. This will give you a maxi- Bradley S. Foster, MD, Springfield, MA Retina & Vitreous mum of 24 credits for the 2008 subscription year. CME credit may be claimed for up to three (3) years from Anil D. Patel, MD, Oklahoma City, OK date of issue. Non-Academy members: For assistance Neuro-Ophthalmology please send an e-mail to [email protected] or a Eric P. Purdy, MD, Fort Wayne, IN fax to (415) 561-8575. Oculoplastic, Lacrimal, & Orbital Surgery Steven I. Rosenfeld, MD, FACS, Delray Beach, FL Refractive Surgery, Optics & Refraction C. Gail Summers, MD, Minneapolis, MN Focal Points (ISSN 0891-8260) is published quarterly by the American Academy of Ophthalmology at 655 Beach St., San Francisco, CA 94109-1336. -

Infectious Keratitis: Short Answers
Q Infectious keratitis: Short answers What is the #1 bacterium in CL-related K ulcer? A Infectious keratitis: Short answers What is the #1 bacterium in CL-related K ulcer? Pseudomonas Infectious keratitis: Short answers Pseudomonas corneal ulcer associated with CL wear Q Infectious keratitis: Short answers What is the #1 bacterium in CL-related K ulcer? Pseudomonas What is the #1 risk factor for Acanthamoeba keratitis? A Infectious keratitis: Short answers What is the #1 bacterium in CL-related K ulcer? Pseudomonas What is the #1 risk factor for Acanthamoeba keratitis? CL wear Infectious keratitis: Short answers Acanthamoeba keratitis associated with CL wear Q Infectious keratitis: Short answers What is the #1 bacterium in CL-related K ulcer? Pseudomonas What is the #1 risk factor for Acanthamoeba keratitis? CL wear What are the three main culprits in fungal keratitis? What is the topical antifungal of choice for each? Fusarium:fungus 1 Topical…natamycin Aspergillisfungus 2 and Candida:fungus 3 A Infectious keratitis: Short answers What is the #1 bacterium in CL-related K ulcer? Pseudomonas What is the #1 risk factor for Acanthamoeba keratitis? CL wear What are the three main culprits in fungal keratitis? What is the topical antifungal of choice for each? Candida: Topical…natamycin Aspergillis and Fusarium: Q Infectious keratitis: Short answers What is the #1 bacterium in CL-related K ulcer? Pseudomonas What is the #1 risk factor for Acanthamoeba keratitis? CL wear What are the three main culprits in fungal keratitis? -

Acanthamoeba, Fungal, and Bacterial Keratitis: a Comparison of Risk Factors and Clinical Features
Acanthamoeba, Fungal, and Bacterial Keratitis: A Comparison of Risk Factors and Clinical Features JEENA MASCARENHAS, PRAJNA LALITHA, N. VENKATESH PRAJNA, MUTHIAH SRINIVASAN, MANORANJAN DAS, SEAN S. D’SILVA, CATHERINE E. OLDENBURG, DURGA S. BORKAR, ELIZABETH J. ESTERBERG, THOMAS M. LIETMAN, AND JEREMY D. KEENAN PURPOSE: To determine risk factors and clinical signs acanthamoeba keratitis research has been conducted in that may differentiate between bacterial, fungal, and industrialized countries, acanthamoeba keratitis also acanthamoeba keratitis among patients presenting with occurs in developing countries, often in non–contact presumed infectious keratitis. lens–wearing individuals.6,7 DESIGN: Hospital-based cross-sectional study. Acanthamoeba keratitis is frequently misdiagnosed as METHODS: We examined the medical records of 115 herpetic or fungal keratitis, and is subsequently treated incor- patients with laboratory-proven bacterial keratitis, 115 rectly, which can lead to poor outcomes.8 Case series of acan- patients with laboratory-proven fungal keratitis, and thamoeba keratitis have identified several important clinical 115 patients with laboratory-proven acanthamoeba kera- signs, such as pseudodendrites, perineural infiltrates, and ring titis seen at Aravind Eye Hospital, Madurai, India, from infiltrates.9,10 However, we are unaware of any studies that 2006-2011. Risk factors and clinical features of the 3 have compared the clinical findings of acanthamoeba organisms were compared using multinomial logistic keratitis with those of bacterial and fungal keratitis. regression. Clinical signs can be especially useful for differentiating the RESULTS: Of 95 patients with bacterial keratitis, 103 cause of infectious keratitis when microbiological testing is patients with fungal keratitis, and 93 patients with acan- not available—which is frequently the case in developing thamoeba keratitis who had medical records available for countries. -

Age, Survival Predictors, and Metastatic Death in Patients with Choroidal Melanoma Tentative Evidence of a Therapeutic Effect on Survival
Research Original Investigation | CLINICAL SCIENCES Age, Survival Predictors, and Metastatic Death in Patients With Choroidal Melanoma Tentative Evidence of a Therapeutic Effect on Survival Bertil E. Damato, MD, PhD, FRCOphth; Heinrich Heimann, MD, FRCOphth; Helen Kalirai, PhD; Sarah E. Coupland, PhD, FRCPath Editorial page 519 IMPORTANCE The influence of ocular treatment of choroidal melanoma on survival has yet to be elucidated. OBJECTIVE To determine whether treatment of choroidal melanoma influences survival by correlating age at death, cause of death, age at treatment, and survival predictors. DESIGN, SETTING, AND PARTICIPANTS Prospective cohort study performed at the Liverpool Ocular Oncology Centre, a supraregional, tertiary referral service in England. We included 3072 patients treated for choroidal melanoma from January 15, 1993, through November 23, 2012, and who reside in the mainland United Kingdom. EXPOSURES A diagnosis of choroidal melanoma (ie, any uveal melanoma involving the choroid). MAIN OUTCOMES AND MEASURES Largest basal tumor diameter, tumor thickness, TNM stage, ciliary body involvement, extraocular spread, melanoma cytomorphological findings, closed connective tissue loops, mitotic count, chromosome 3 loss, chromosome 6p gain, chromosome 8q gain, age at treatment, age at death, and cause of death. RESULTS The largest basal tumor diameter correlated with all survival predictors except for chromosome 6p gain. Older age at treatment correlated with ciliary body involvement, extraocular spread, largest basal tumor diameter, tumor thickness, TNM stage, epithelioid cells, chromosome 3 loss, and chromosome 8q gain. A total of 1005 patients had died by the close of the study. The cause of death was metastatic disease due to uveal melanoma in 561 patients. Among the 561 patients, survival time after treatment correlated with sex, basal tumor diameter, ciliary body involvement, extraocular spread, TNM stage, closed loops, and mitotic count. -

Ophthalmic Drugs
A Supplement to 22nd EDITION Randall Thomas, OD, MPH Patrick Vollmer, OD The Clinical Guide to Dr. Melton Ophthalmic[ [ Dr. Thomas Drugs Dispense as writ ten — no substit utions. Refills: unlimit ed. May 15, 2018 Dr. Vollmer Peer-to-peer advice to help boost your prescribing prowess. Supported by an unrestricted grant from Bausch + Lomb 001_dg0518_fc.indd 3 5/11/18 10:52 AM FROM THE AUTHORS DEAR OPTOMETRIC COLLEAGUES: Supported by an Welcome to the 2018 edition of our annual Clinical Guide to Ophthalmic unrestricted grant from Drugs. Herein, we provide updates on our collective clinical experiences and Bausch + Lomb heavily season them with pertinent excerpts from the literature. This guide is intended to bring solid, scientifically accurate and clinically relevant information to our optometric colleagues. If you want to understand CONTENTS how the three of us treat, and what factors led us to develop these methods, you’ll find it explained here. The methods and opinions represented are our own. We recognize that other doctors may use alternative approaches. That First-year Impressions ...........3 is true in all of health care. But this three-doctor writing team has logged over 75 combined years of clinical optometry, and we bring that ‘real-world’ spirit to the discussions that follow. Know that, above all, we are doctors who are genuinely concerned for our patients’ well-being and who endeavor to Glaucoma Care .......................... 6 provide them the best of care, and we write from that perspective. The two topics of greatest interest and need for most eye physicians right now are glaucoma and dry eye disease.