Genomic Dynamics of Transposable Elements in the Western Clawed Frog (Silurana Tropicalis)
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Genome Sequence of the Progenitor of the Wheat D Genome Aegilops Tauschii Ming-Cheng Luo1*, Yong Q
OPEN LETTER doi:10.1038/nature24486 Genome sequence of the progenitor of the wheat D genome Aegilops tauschii Ming-Cheng Luo1*, Yong Q. Gu2*, Daniela Puiu3*, Hao Wang4,5,6*, Sven O. Twardziok7*, Karin R. Deal1, Naxin Huo1,2, Tingting Zhu1, Le Wang1, Yi Wang1,2, Patrick E. McGuire1, Shuyang Liu1, Hai Long1, Ramesh K. Ramasamy1, Juan C. Rodriguez1, Sonny L. Van1, Luxia Yuan1, Zhenzhong Wang1,8, Zhiqiang Xia1, Lichan Xiao1, Olin D. Anderson2, Shuhong Ouyang2,8, Yong Liang2,8, Aleksey V. Zimin3, Geo Pertea3, Peng Qi4,5, Jeffrey L. Bennetzen6, Xiongtao Dai9, Matthew W. Dawson9, Hans-Georg Müller9, Karl Kugler7, Lorena Rivarola-Duarte7, Manuel Spannagl7, Klaus F. X. Mayer7,10, Fu-Hao Lu11, Michael W. Bevan11, Philippe Leroy12, Pingchuan Li13, Frank M. You13, Qixin Sun8, Zhiyong Liu8, Eric Lyons14, Thomas Wicker15, Steven L. Salzberg3,16, Katrien M. Devos4,5 & Jan Dvořák1 Aegilops tauschii is the diploid progenitor of the D genome of We conclude therefore that the size of the Ae. tauschii genome is about hexaploid wheat1 (Triticum aestivum, genomes AABBDD) and 4.3 Gb. an important genetic resource for wheat2–4. The large size and To assess the accuracy of our assembly, sequences of 195 inde- highly repetitive nature of the Ae. tauschii genome has until now pendently sequenced and assembled AL8/78 BAC clones8, which precluded the development of a reference-quality genome sequence5. contained 25,540,177 bp in 2,405 unordered contigs, were aligned to Here we use an array of advanced technologies, including ordered- Aet v3.0. Five contigs failed to align and six extended partly into gaps, clone genome sequencing, whole-genome shotgun sequencing, accounting for 0.25% of the total length of the contigs. -

Genomes:Genomes: Whatwhat Wewe Knowknow …… Andand Whatwhat Wewe Don’Tdon’T Knowknow
Genomes:Genomes: WhatWhat wewe knowknow …… andand whatwhat wewe don’tdon’t knowknow Complete draft sequence 2001 OctoberOctober 15,15, 20072007 Dr.Dr. StefanStefan Maas,Maas, BioSBioS Lehigh Lehigh U.U. © SMaas 2007 What we know Raw genome data © SMaas 2007 The range of genome sizes in the animal & plant kingdoms !! NoNo correlationcorrelation betweenbetween genomegenome sizesize andand complexitycomplexity © SMaas 2007 What accounts for the often massive and seemingly arbitrary differences in genome size observed among eukaryotic organisms? The fruit fly The mountain grasshopper Drosophila melanogaster Podisma pedestris 180 Mb 18,000 Mb The difference in genome size of a factor of 100 is difficult to explain in view of the apparently similar levels of evolutionary, developmental and behavioral complexity of these organisms. © SMaas 2007 ComplexityComplexity doesdoes notnot correlatecorrelate withwith genomegenome sizesize 3.4 × 10 9 bp 6.7 × 1011 bp Homo sapiens Amoeba dubia © SMaas 2007 ComplexityComplexity doesdoes notnot correlatecorrelate withwith genegene numbernumber ~31,000~31,000 genesgenes ~26,000~26,000 genesgenes ~50,000~50,000 genesgenes © SMaas 2007 IsIs anan ExpansionExpansion inin GeneGene NumberNumber drivingdriving EvolutionEvolution ofof HigherHigher Organisms?Organisms? Vertebrata 30,000 Urochordata 16,000 Arthropoda 14,000 Nematoda 21,000 Fungi 2,000 – 13,000 Vascular plants 25,000 – 60,000 Unicellular sps. 5,000 – 10,000 Prokaryotes 500 - 7,000 © SMaas 2007 Structure of DNA Watson and Crick in 1953 proposed that DNA is a double helix in which the 4 bases are base paired, Adenine (A) with Thymine (T) and Guanine (G) with Cytosine (C). © SMaas 2007 © SMaas 2007 © SMaas 2007 Steps in the folding of DNA to create an eukaryotic chromosome 30 nm fiber (6 nucleosomes per turn) FactorFactor ofof condensation:condensation: Ca.Ca. -

Inbreeding Ratio and Genetic Relationships Among Strains of the Western Clawed Frog, Xenopus Tropicalis
RESEARCH ARTICLE Inbreeding Ratio and Genetic Relationships among Strains of the Western Clawed Frog, Xenopus tropicalis Takeshi Igawa1¤*, Ai Watanabe1, Atsushi Suzuki1, Akihiko Kashiwagi1, Keiko Kashiwagi1, Anna Noble2, Matt Guille2, David E. Simpson3, Marko E. Horb4, Tamotsu Fujii5, Masayuki Sumida1 1 Institute for Amphibian Biology, Graduate School of Science, Hiroshima University, Higashi-Hiroshima, Hiroshima, Japan, 2 School of Biological Sciences, Institute of Biomedical and Biomolecular Science, University of Portsmouth, Portsmouth, United Kingdom, 3 The Wellcome Trust/Cancer Research UK Gurdon Institute, The Henry Wellcome Building of Cancer and Developmental Biology, University of Cambridge, Cambridge, United Kingdom, 4 Bell Center for Regenerative Biology and Tissue Engineering and National Xenopus Resource, Marine Biological Laboratory, Woods Hole, MA, United States of America, 5 Department of Health Sciences, Faculty of Human Culture & Science, Prefectural University of Hiroshima, Hiroshima, Japan ¤ Current Address: Division of Development Science, Graduate School of International Development and OPEN ACCESS Cooperation, Hiroshima University, Higashi-Hiroshima, Hiroshima, Japan * [email protected] Citation: Igawa T, Watanabe A, Suzuki A, Kashiwagi A, Kashiwagi K, Noble A, et al. (2015) Inbreeding Ratio and Genetic Relationships among Strains of the Western Clawed Frog, Xenopus tropicalis. PLoS Abstract ONE 10(7): e0133963. doi:10.1371/journal. pone.0133963 The Western clawed frog, Xenopus tropicalis, is a highly promising model amphibian, espe- Editor: Gert Jan C. Veenstra, Radboud University cially in developmental and physiological research, and as a tool for understanding disease. Nijmegen, NETHERLANDS It was originally found in the West African rainforest belt, and was introduced to the research Received: January 8, 2015 community in the 1990s. -

Saccharomyces Cerevisiae
letters to nature 19Department of Yeast Genetics, Institute of Molecular Medecine, John Radcliffe Hospital, Headington, Oxford OX3 9DU, UK The nucleotide sequence of 20L.N.C.I.B., Area Science Park, Padriciano 99, I-34012 Trieste, Italy 21GATC GmbH, Fitz-Arnold-Strasse 23, 78467 Konstanz, Germany Saccharomyces cerevisiae 22Carlsberg Laboratory, Gamle Carlsberg Vej 10, DK-2500 Copenhagen Valby, Denmark chromosome IV 23AGON GmbH, Glienicker Weg 185, D-12489 Berlin, Germany 24Katholieke Universiteit Leuven, Laboratory of Gene Technology, Willem de C. Jacq1, J. Alt-Mörbe2, B. Andre3, W. Arnold4, A. Bahr5, Croylaan, 42, B-3001 Leuven, Belgium J. P. G. Ballesta6, M. Bargues7, L. Baron8, A. Becker4, N. Biteau8, 25The Sanger Centre, Wellcome Trust Genome Campus, Hinxton, Cambridge H. Blöcker9, C. Blugeon1, J. Boskovic6, P. Brandt9, M. Brückner10 , CB10 1SA, UK M. J. Buitrago11 , F. Coster12, T. Delaveau1, F. del Rey11 , B. Dujon13, 26Department of Biochemistry, Stanford University, Beckman Center, Stanford L. G. Eide14, J. M. Garcia-Cantalejo6, A. Goffeau12, A. Gomez-Peris15, CA 94305-5307, USA C. Granotier8, V. Hanemann16, T. Hankeln5, J. D. Hoheisel17, W. Jäger9, 27The Genome Sequencing Center, Department of Genetics, Washington University, A. Jimenez6, J.-L. Jonniaux12, C. Krämer5, H. Küster4, P. Laamanen18, School of Medicine, 630 S. Euclid Avenue, St Louis, Missouri 63110, USA Y. Legros8, E. Louis19, S. Möller-Rieker5, A. Monnet8, M. Moro20, 28Martinsrieder Institut für Protein Sequenzen, Max-Planck-Institut für S. Müller-Auer10 , B. Nußbaumer4, N. Paricio7, L. Paulin18, J. Perea1, Biochemie, D-82152 Martinsried bei München, Germany. M. Perez-Alonso7, J. E. Perez-Ortin15, T. M. Pohl21, H. Prydz14, B. -
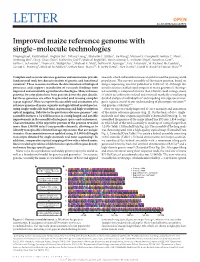
Improved Maize Reference Genome with Single-Molecule Technologies Yinping Jiao1, Paul Peluso2, Jinghua Shi3, Tiffany Liang3, Michelle C
OPEN LETTER doi:10.1038/nature22971 Improved maize reference genome with single-molecule technologies Yinping Jiao1, Paul Peluso2, Jinghua Shi3, Tiffany Liang3, Michelle C. Stitzer4, Bo Wang1, Michael S. Campbell1, Joshua C. Stein1, Xuehong Wei1, Chen-Shan Chin2, Katherine Guill5, Michael Regulski1, Sunita Kumari1, Andrew Olson1, Jonathan Gent6, Kevin L. Schneider7, Thomas K. Wolfgruber7, Michael R. May8, Nathan M. Springer9, Eric Antoniou1, W. Richard McCombie1, Gernot G. Presting7, Michael McMullen5, Jeffrey Ross-Ibarra10, R. Kelly Dawe6, Alex Hastie3, David R. Rank2 & Doreen Ware1,11 Complete and accurate reference genomes and annotations provide research, which will enable increases in yield to feed the growing world fundamental tools for characterization of genetic and functional population. The current assembly of the maize genome, based on variation1. These resources facilitate the determination of biological Sanger sequencing, was first published in 2009 (ref. 3). Although this processes and support translation of research findings into initial reference enabled rapid progress in maize genomics1, the origi- improved and sustainable agricultural technologies. Many reference nal assembly is composed of more than 100,000 small contigs, many genomes for crop plants have been generated over the past decade, of which are arbitrarily ordered and oriented, markedly complicating but these genomes are often fragmented and missing complex detailed analysis of individual loci6 and impeding investigation of inter- repeat regions2. Here we report the assembly and annotation of a genic regions crucial to our understanding of phenotypic variation7,8 reference genome of maize, a genetic and agricultural model species, and genome evolution9,10. using single-molecule real-time sequencing and high-resolution Here we report a vastly improved de novo assembly and annotation optical mapping. -

Apis Mellifera)
COMPUTATIONAL ANALYSIS OF DIFFICULT-TO-PREDICT GENES AND DETECTION OF LINEAGE-SPECIFIC GENES IN HONEY BEE (APIS MELLIFERA) A Dissertation submitted to the Faculty of the Graduate School of Arts and Sciences of Georgetown University in partial fulfillment of the requirements for the degree of Doctor of Philosophy in Biology. By Anna K. Bennett, M.S. Washington, DC July 29, 2013 Copyright 2013 by Anna K. Bennett All Rights Reserved ii COMPUTATIONAL ANALYSIS OF DIFFICULT-TO-PREDICT GENES AND DETECTION OF LINEAGE-SPECIFIC GENES IN HONEY BEE (APIS MELLIFERA) Anna K. Bennett, M.S. Thesis Advisor: Christine G. Elsik, Ph.D. ABSTRACT Honey bees are key agricultural pollinators and a research model for social behavior and the evolution of eusociality. Generation of reliable gene predictions is critical to the success of laboratory experiments and comparative analyses for sequenced genomes. The honey bee genome, published in 2006 by the Honey Bee Genome Sequencing Consortium, had fewer predicted genes than expected, partially due to a lack of transcriptome data and protein homologs from closely related species. As part of ongoing Consortium efforts, genomes of two closely related taxa were sequenced, dwarf honey bee (Apis florea) and buff-tailed bumble bee (Bombus terrestris), the draft A. mellifera genome was improved with additional sequence, and multiple A. mellifera tissue transcriptomes were sequenced. The Consortium predicted genes using seven methods and used these data to produce an improved official gene set (OGSv3.2) with ~5000 more protein-coding genes than the first set (OGSv1.0). We present our approach to detect previously unknown genes in A. -

Genome-Wide Analysis of Admixture and Adaptation in the Africanized Honeybee
Nelson, R. M., Wallberg, A., Simões, Z. L. P., Lawson, D. J., & Webster, M. T. (2017). Genome-wide analysis of admixture and adaptation in the Africanized honeybee. Molecular Ecology, 26(14), 3603-3617. https://doi.org/10.1111/mec.14122 Peer reviewed version License (if available): CC BY-NC Link to published version (if available): 10.1111/mec.14122 Link to publication record in Explore Bristol Research PDF-document This is the author accepted manuscript (AAM). The final published version (version of record) is available online via Wiley at https://onlinelibrary.wiley.com/doi/abs/10.1111/mec.14122 . Please refer to any applicable terms of use of the publisher. University of Bristol - Explore Bristol Research General rights This document is made available in accordance with publisher policies. Please cite only the published version using the reference above. Full terms of use are available: http://www.bristol.ac.uk/red/research-policy/pure/user-guides/ebr-terms/ Genome-wide analysis of admixture and adaptation in the Africanized honeybee Ronald M. Nelson1, Andreas Wallberg1, Zilá Luz Paulino Simões2, Daniel J. Lawson3, Matthew T. Webster1* 1. Department of Medical Biochemistry and Microbiology, Science for Life Laboratory, Uppsala University, Uppsala, Sweden. 2. Department of Biology, University of São Paulo, São Paulo, Brazil. 3. Department of Mathematics, University of Bristol, Bristol, United Kingdom. Keywords: Africanized honeybee, admixture, introgression, adaptation, biological invasion, natural selection *[email protected] Running title: Adaptation in Africanized honeybees 1 Abstract Genetic exchange by hybridization or admixture can make an important contribution to evolution, and introgression of favourable alleles can facilitate adaptation to new environments. -

Frogs Reveal Mechanism That Determines Viability of Hybrids 11 January 2018
Frogs reveal mechanism that determines viability of hybrids 11 January 2018 which one is the mother. One hybrid is viable, the other hybrid is not Both mules and hinnies are infertile, because donkeys and horses have a different number of chromosomes, the protein structures on which Crossbreeding Xenopus laevis, the African clawed frog, genetic material is present. Yeasts, plants, fish and and Xenopus tropicalis, the Western clawed frog has amphibians (in contrast to mammals) can, however, asymmetric results. When a female African clawed frog produce fertile hybrids. Professor of Molecular is crossbred with a male Western clawed frog, the Developmental Biology Gert Jan Veenstra says, embryos are viable. However, the other way around, "For instance, in frogs a duplication of crossbreeding a male African clawed frog and a female Western clawed frog leads to embryos that die in the chromosomes can appear, in which the whole set early stages of development. Researchers found the of chromosomes from the father and the mother are reason for this: the maternal molecular machinery of the passed on to the next generation." Western clawed frog cannot fully recognize the paternal chromosomes of the African clawed frog. The separation However, there is another problem: Some hybrids of the paternal chromosomes during cell division is are not viable, while crossbreeding the other way disrupted. As a consequence, the cells lack a large around is. The embryos are genetically identical, number of vital genes and quickly die. Credit: University but there is a difference in viability, depending on of California Berkeley, Radboud University the father species and the mother species. -

Recombination in Diverse Maize Is Stable, Predictable, and Associated with Genetic Load
Recombination in diverse maize is stable, predictable, and associated with genetic load Eli Rodgers-Melnicka,1, Peter J. Bradburya,b,1, Robert J. Elshirea, Jeffrey C. Glaubitza, Charlotte B. Acharyaa, Sharon E. Mitchella, Chunhui Lic, Yongxiang Lic, and Edward S. Bucklera,b aInstitute for Genomic Diversity, Cornell University, Ithaca, NY 14853; bUS Department of Agriculture-Agricultural Research Service, Ithaca, NY 14853; and cInstitute of Crop Science, Chinese Academy of Agricultural Sciences, Beijing 100081, China Edited by Qifa Zhang, Huazhong Agricultural University, Wuhan, China, and approved February 6, 2015 (received for review July 21, 2014) Among the fundamental evolutionary forces, recombination ar- On a molecular level, chromatin structure heavily influences guably has the largest impact on the practical work of plant the cross-over rate in plants. Not only are heterochromatic breeders. Varying over 1,000-fold across the maize genome, the regions generally depleted of cross-overs (11), but KO of cytosine- local meiotic recombination rate limits the resolving power of DNA-methyl-transferase (MET1)inArabidopsis thaliana leads quantitative trait mapping and the precision of favorable allele to both genome-wide CpG hypomethylation and a relative in- introgression. The consequences of low recombination also theo- crease in the proportion of cross-overs within the euchromatic – retically extend to the species-wide scale by decreasing the power chromosomal arms (12 14). Nucleotide content may also be as- of selection relative to genetic drift, and thereby hindering the sociated with the local frequency of recombination, potentially purging of deleterious mutations. In this study, we used genotyp- due to the effect of GC-biased gene conversion (bGC) during res- ing-by-sequencing (GBS) to identify 136,000 recombination break- olution of heteroduplexes that form at cross-over junctions (15). -

Co-Expression of Neighboring Genes in the Zebrafish (Danio Rerio) Genome
Int. J. Mol. Sci. 2009, 10, 3658-3670; doi:10.3390/ijms10083658 OPEN ACCESS International Journal of Molecular Sciences ISSN 1422-0067 www.mdpi.com/journal/ijms Article Co-Expression of Neighboring Genes in the Zebrafish (Danio rerio) Genome Huai-Kuang Tsai 1, Pei-Ying Huang 2, Cheng-Yan Kao 2 and Daryi Wang 3,* 1 Institute of Information Science, Academia Sinica, 128 Sec. 2, Academia Rd, Nankang, 115, Taipei, Taiwan; E-Mail: [email protected] (H.-K.T.) 2 Department of Computer Science and Information Engineering, National Taiwan University, Taipei 106, Taiwan; E-Mails: [email protected] (P.-Y.H.); [email protected] (C.-Y.K.) 3 Biodiversity Research Center, Academia Sinica, 128 Sec. 2, Academia Rd, Nankang, 115, Taipei, Taiwan * Author to whom correspondence should be addressed; E-Mail: [email protected] (D.W.); Tel. +886-2-27890159; Fax: +886-2-27829624. Received: 14 July 2009; in revised form: 11 August 2009 / Accepted: 20 August 2009 / Published: 21 August 2009 Abstract: Neighboring genes in the eukaryotic genome have a tendency to express concurrently, and the proximity of two adjacent genes is often considered a possible explanation for their co-expression behavior. However, the actual contribution of the physical distance between two genes to their co-expression behavior has yet to be defined. To further investigate this issue, we studied the co-expression of neighboring genes in zebrafish, which has a compact genome and has experienced a whole genome duplication event. Our analysis shows that the proportion of highly co-expressed neighboring pairs (Pearson’s correlation coefficient R>0.7) is low (0.24% ~ 0.67%); however, it is still significantly higher than that of random pairs. -

Ceranae, an Emergent Pathogen of Honey Bees
Genomic Analyses of the Microsporidian Nosema ceranae, an Emergent Pathogen of Honey Bees R. Scott Cornman1, Yan Ping Chen1, Michael C. Schatz2, Craig Street3, Yan Zhao4, Brian Desany5, Michael Egholm5, Stephen Hutchison5, Jeffery S. Pettis1, W. Ian Lipkin3, Jay D. Evans1* 1 USDA-ARS Bee Research Lab, Beltsville, Maryland, United States of America, 2 Center for Bioinformatics and Computational Biology, University of Maryland, College Park, Maryland, United States of America, 3 Center for Infection and Immunity, Mailman School of Public Health, Columbia University, New York, New York, United States of America, 4 USDA-ARS Molecular Plant Pathology Laboratory, Beltsville, Maryland, United States of America, 5 454 Life Sciences/Roche Applied Sciences, Branford, Connecticut, United States of America Abstract Recent steep declines in honey bee health have severely impacted the beekeeping industry, presenting new risks for agricultural commodities that depend on insect pollination. Honey bee declines could reflect increased pressures from parasites and pathogens. The incidence of the microsporidian pathogen Nosema ceranae has increased significantly in the past decade. Here we present a draft assembly (7.86 MB) of the N. ceranae genome derived from pyrosequence data, including initial gene models and genomic comparisons with other members of this highly derived fungal lineage. N. ceranae has a strongly AT-biased genome (74% A+T) and a diversity of repetitive elements, complicating the assembly. Of 2,614 predicted protein-coding sequences, we conservatively estimate that 1,366 have homologs in the microsporidian Encephalitozoon cuniculi, the most closely related published genome sequence. We identify genes conserved among microsporidia that lack clear homology outside this group, which are of special interest as potential virulence factors in this group of obligate parasites. -

Efficient Targeted Gene Disruption in the Soma and Germ Line of the Frog
Efficient targeted gene disruption in the soma and germ line of the frog Xenopus tropicalis using engineered zinc-finger nucleases John J. Younga, Jennifer M. Cheroneb, Yannick Doyonb, Irina Ankoudinovab, Farhoud M. Farajib, Andrew H. Leeb, Catherine Ngob, Dmitry Y. Guschinb, David E. Paschonb, Jeffrey C. Millerb, Lei Zhangb, Edward J. Rebarb, Philip D. Gregoryb, Fyodor D. Urnovb, Richard M. Harlanda,1, and Bryan Zeitlerb aDepartment of Molecular and Cell Biology, University of California, Berkeley, CA 94720-3200; and bSangamo BioSciences, Inc., Richmond, CA 94804 Edited* by John Gerhart, University of California, Berkeley, CA, and approved March 10, 2011 (received for review February 7, 2011) The frog Xenopus, an important research organism in cell and de- To develop conditions for gene disruption in X. tropicalis,we velopmental biology, currently lacks tools for targeted mutagen- made use of transgenic animals carrying a single-copy GFP esis. Here, we address this problem by genome editing with zinc- transgene (14). Wild-type X. tropicalis eggs were fertilized with finger nucleases (ZFNs). ZFNs directed against an eGFP transgene in sperm from a homozygous GFP transgenic male. The resulting Xenopus tropicalis induced mutations consistent with nonhomol- heterozygous embryos were injected with mRNA-encoding ZFNs ogous end joining at the target site, resulting in mosaic loss of the that target the eGFP coding region (15). Uninjected tadpoles ex- fluorescence phenotype at high frequencies. ZFNs directed against press GFP robustly in the somites, lens, and head musculature the noggin gene produced tadpoles and adult animals carrying up (Fig. 1 A and B). Injection of 20 pg eGFP ZFN RNAs led to mosaic to 47% disrupted alleles, and founder animals yielded progeny loss of fluorescence in otherwise healthy tadpoles (Fig.