Two Grizzly Bear Studies : Moth Feeding Ecology and Male
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Field Notes in Support of a Revision of Hawaiian Carabidae
Field Notes in Support of a Revision of Hawaiian Carabidae James K. Liebherr* Cornell University Ithaca, NY, U.S.A. * Curator (Ret.) Cornell University Insect Collection, and Professor Emeritus 23 August 2021 Field Notes in Support of a Revision of Hawaiian Carabidae © 2021 by James K. Liebherr is licensed under Attribution-NonCommercial-NoDerivatives 4.0 International. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-nd/4.0/ Cornell University Insect Collection John H. and Ann B. Comstock Hall Cornell University Ithaca, NY 14853-2601 U.S.A. ii Table of Contents Foreword ……………………………………………………………….…………………………. iv 00.—Introduction …………………………………………………………………..………….. 1 01.—Kauaˋi; 9-22 March, 10-16 May 1991 ……..…………………………...….... 7 02.—Hawaiˋi Island; 23 March-5 April 1991 ..…………………………..…….… 21 03.—Haleakalā; 8-13 April 1991, 29 April–9 May 1991 …………….…….… 27 04.—Oˋahu; 20–23 April, 18 May 1991 ………………………..……….…..….…..41 05.—Molokaˋi; 24-27 April 1991 …………………………………….……..….………45 06.—West Maui, 11-15 May 1992; Molokaˋi, 15-16 May 1992 ...……… 50 07.—Maui Nui, 29 April-21 May 1993 (Lānaˋi, Molokaˋi, West Maui, Haleakalā) ……………….………………………………………………………………….…….61 08.—Oˋahu, 1-15 May 1995 .………………………………….……………………..…. 89 09.—Kauaˋi, 16-22 May 1995 ………………….………………..…………..………. 104 10.—Maui + Molokaˋi 20-31 May 1997 …..……………………………………... 114 11.—Hawaiˋi Island—1–6 June 1997 …..………………………..……………….. 134 12.—Hawaiˋi Island—9–17 October 1997 …..……………………….…….….. 138 13.—Haleakalā, Maui—30 April–15 May 1998 ….…………………………... 145 14.—Haleakalā and West Maui—15–23 May 2001 …….………….………. 167 15.—Hawaiˋi Island—23–27 May 2001 …………………..…………..…………. 179 16.—Haleakalā, Maui, 14–20 May 2003 …………………………………………. 184 17.—Hawaiˋi Island, 21–25 May 2003 ………………..………………………….. 192 18.—Lānaˋi, Molokaˋi, West Maui, 10–28 May 2004 …..……..….………. -

BIOLOGICAL CONTROL of the BLACK CUTWORM, <Em>AGROTIS IPSILON</Em> (LEPIDOPTERA: NOCTUIDAE), and ENDOPHYTE MEDIATED T
University of Kentucky UKnowledge University of Kentucky Doctoral Dissertations Graduate School 2011 BIOLOGICAL CONTROL OF THE BLACK CUTWORM, AGROTIS IPSILON (LEPIDOPTERA: NOCTUIDAE), AND ENDOPHYTE MEDIATED TRITROPHIC INTERACTIONS IN TURFGRASS Andrea Jeanne Bixby-Brosi University of Kentucky, [email protected] Right click to open a feedback form in a new tab to let us know how this document benefits ou.y Recommended Citation Bixby-Brosi, Andrea Jeanne, "BIOLOGICAL CONTROL OF THE BLACK CUTWORM, AGROTIS IPSILON (LEPIDOPTERA: NOCTUIDAE), AND ENDOPHYTE MEDIATED TRITROPHIC INTERACTIONS IN TURFGRASS" (2011). University of Kentucky Doctoral Dissertations. 183. https://uknowledge.uky.edu/gradschool_diss/183 This Dissertation is brought to you for free and open access by the Graduate School at UKnowledge. It has been accepted for inclusion in University of Kentucky Doctoral Dissertations by an authorized administrator of UKnowledge. For more information, please contact [email protected]. ABSTRACT OF DISSERTATION Andrea Jeanne Bixby-Brosi The Graduate School University of Kentucky 2011 BIOLOGICAL CONTROL OF THE BLACK CUTWORM, AGROTIS IPSILON (LEPIDOPTERA: NOCTUIDAE), AND ENDOPHYTE MEDIATED TRITROPHIC INTERACTIONS IN TURFGRASS ABSTRACT OF DISSERTATION A dissertation submitted in partial fulfillment of the requirements for the degree of Doctor of Philosophy in the College of Agriculture at the University of Kentucky By Andrea Jeanne Bixby-Brosi Lexington, Kentucky Director: Dr. Daniel A. Potter, Professor of Entomology Lexington, Kentucky 2011 Copyright © Andrea Jeanne Bixby-Brosi 2011 ABSTRACT OF DISSERTATION BIOLOGICAL CONTROL OF THE BLACK CUTWORM, AGROTIS IPSILON (LEPIDOPTERA: NOCTUIDAE), AND ENDOPHYTE MEDIATED TRITROPHIC INTERACTIONS IN TURFGRASS Components of successful pest management programs must be complementary and not antagonistic. This project examined interactions between natural enemies of the black cutworm, Agrotis ipsilon (Hufnagel), an important turfgrass pest, and host plant resistance by endophytic grass. -

Agrotis Ipsilon): a Glimpse on Globally Important Crop Pest
Volume 2021, issue 2, pages 36–42 30 June 2021 https://doi.org/10.33493/scivis.21.02.02 REVIEW ARTICLE An insight into black cutworm (Agrotis ipsilon): A glimpse on globally important crop pest Christopher Rodingpuia*, H. Lalthanzara Department of Life Sciences, Pachhunga University College, Mizoram University, Aizawl 796001, India Many agricultural pests create problems for farmers leading to production loss. Received 11 January 2021 Accepted 15 February 2021 Since agriculture plays an important role in the economy of a nation, controlling and management of such pests is of great importance. Cutworms are notorious *For correspondence: [email protected] agricultural pests infesting a wide variety of crops leading to production loss. Cutworms are distributed worldwide and many species of cutworms have also Contact us: been reported feeding on all kinds of crops. Agrotis ipsilon is one of the most [email protected] common cutworm species prevailing in different continents. Understanding the biology and management of these pests will be of great use for farmers. In this article, brief information on cutworms; specifically A. ipsilon species is highlighted. A brief description of host, migration, control and management of A. ipsilon are also highlighted. Keywords: Caterpillars, infestation, host, parasite, economy, control-management. Introduction Cutworms are caterpillars belonging to the order may extend up to 1 inch (2.5 cm).4 There are Lepidoptera and family Noctuidae; a large family of variations within the genera and may differ moth.1-4 Cutworms got their names from their depending on the region where they are found.1-4 feeding habit of “cutting-off”’ a seedling at ground Most common cutworm species are Agrotis level by chewing through the stem. -
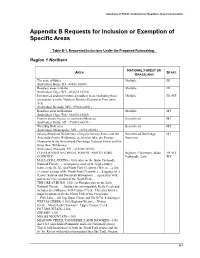
Summary of Public Comment, Appendix B
Summary of Public Comment on Roadless Area Conservation Appendix B Requests for Inclusion or Exemption of Specific Areas Table B-1. Requested Inclusions Under the Proposed Rulemaking. Region 1 Northern NATIONAL FOREST OR AREA STATE GRASSLAND The state of Idaho Multiple ID (Individual, Boise, ID - #6033.10200) Roadless areas in Idaho Multiple ID (Individual, Olga, WA - #16638.10110) Inventoried and uninventoried roadless areas (including those Multiple ID, MT encompassed in the Northern Rockies Ecosystem Protection Act) (Individual, Bemidji, MN - #7964.64351) Roadless areas in Montana Multiple MT (Individual, Olga, WA - #16638.10110) Pioneer Scenic Byway in southwest Montana Beaverhead MT (Individual, Butte, MT - #50515.64351) West Big Hole area Beaverhead MT (Individual, Minneapolis, MN - #2892.83000) Selway-Bitterroot Wilderness, along the Selway River, and the Beaverhead-Deerlodge, MT Anaconda-Pintler Wilderness, at Johnson lake, the Pioneer Bitterroot Mountains in the Beaverhead-Deerlodge National Forest and the Great Bear Wilderness (Individual, Missoula, MT - #16940.90200) CLEARWATER NATIONAL FOREST: NORTH FORK Bighorn, Clearwater, Idaho ID, MT, COUNTRY- Panhandle, Lolo WY MALLARD-LARKINS--1300 (also on the Idaho Panhandle National Forest)….encompasses most of the high country between the St. Joe and North Fork Clearwater Rivers….a low elevation section of the North Fork Clearwater….Logging sales (Lower Salmon and Dworshak Blowdown) …a potential wild and scenic river section of the North Fork... THE GREAT BURN--1301 (or Hoodoo also on the Lolo National Forest) … harbors the incomparable Kelly Creek and includes its confluence with Cayuse Creek. This area forms a major headwaters for the North Fork of the Clearwater. …Fish Lake… the Jap, Siam, Goose and Shell Creek drainages WEITAS CREEK--1306 (Bighorn-Weitas)…Weitas Creek…North Fork Clearwater. -

Region Forest Roadless Name GIS Acres 1 Beaverhead-Deerlodge
These acres were calculated from GIS data Available on the Forest Service Roadless website for the 2001 Roadless EIS. The data was downloaded on 8/24/2011 by Suzanne Johnson WO Minerals & Geology‐ GIS/Database Specialist. It was discovered that the Santa Fe NF in NM has errors. This spreadsheet holds the corrected data from the Santa Fe NF. The GIS data was downloaded from the eGIS data center SDE instance on 8/25/2011 Region Forest Roadless Name GIS Acres 1 Beaverhead‐Deerlodge Anderson Mountain 31,500.98 1 Beaverhead‐Deerlodge Basin Creek 9,499.51 1 Beaverhead‐Deerlodge Bear Creek 8,122.88 1 Beaverhead‐Deerlodge Beaver Lake 11,862.81 1 Beaverhead‐Deerlodge Big Horn Mountain 50,845.85 1 Beaverhead‐Deerlodge Black Butte 39,160.06 1 Beaverhead‐Deerlodge Call Mountain 8,795.54 1 Beaverhead‐Deerlodge Cattle Gulch 19,390.45 1 Beaverhead‐Deerlodge Cherry Lakes 19,945.49 1 Beaverhead‐Deerlodge Dixon Mountain 3,674.46 1 Beaverhead‐Deerlodge East Pioneer 145,082.05 1 Beaverhead‐Deerlodge Electric Peak 17,997.26 1 Beaverhead‐Deerlodge Emerine 14,282.26 1 Beaverhead‐Deerlodge Fleecer 31,585.50 1 Beaverhead‐Deerlodge Flint Range / Dolus Lakes 59,213.30 1 Beaverhead‐Deerlodge Four Eyes Canyon 7,029.38 1 Beaverhead‐Deerlodge Fred Burr 5,814.01 1 Beaverhead‐Deerlodge Freezeout Mountain 97,304.68 1 Beaverhead‐Deerlodge Garfield Mountain 41,891.22 1 Beaverhead‐Deerlodge Goat Mountain 9,347.87 1 Beaverhead‐Deerlodge Granulated Mountain 14,950.11 1 Beaverhead‐Deerlodge Highlands 20,043.87 1 Beaverhead‐Deerlodge Italian Peak 90,401.31 1 Beaverhead‐Deerlodge Lone Butte 13,725.16 1 Beaverhead‐Deerlodge Mckenzie Canyon 33,350.48 1 Beaverhead‐Deerlodge Middle Mtn. -

Aka "Miller Moth"
Questions and Answers about Miller Moths by Whitney Cranshaw and Frank Peairs Colorado State University Extension Entomologists Prediction of Nuisance Problems with Army Cutworm (a.k.a. "Miller Moth") for April 25, 2014 There have been several reports that the larvae of the army cutworm are unusually abundant this spring on the plains of northeastern Colorado and adjacent areas of Kansas and Nebraska. This poses risk to infested crops, and some treatments are being made in wheat and alfalfa. However, army cutworm numbers are also a good predictor of the population size of the adults (“miller moths”) to be expected as these insects complete development and make their annual migration to the higher elevations. Based on the stage of cutworm larvae present, adults likely will begin to appear in about a month and a half, but the peak of the migration will not pass through the northern Front Range until a bit later (end of May, early June). The numbers of caterpillars present suggest that the numbers of adults migrating across northeastern Colorado will be well above average. Drought conditions that persist in southeastern Colorado apparently have decreased populations of army cutworms in that part of the state. There are no indications that army cutworms, and the adult “miller moths”, originating out of southeastern Colorado will be unusually abundant this year; an average, perhaps somewhat below average, number of “miller moths” may be seen in that part of the state. However, those that are present will likely concentrate in irrigated areas where there are blooming plants in late May and June. -

Peak Bagging
Peak Bagging: (complete award size: 8" x 6") Program and Awards Offered by the HIGH ADVENTURE TEAM Greater Los Angeles Area Council Boy Scouts of America The High Adventure Team of the Greater Los Angeles Area Council-Boy Scouts of America is a volunteer group of Scouters which operates under the direction of GLAAC-Camping Services. Its mission is to develop and promote outdoor activities within the Council and by its many Units. It conducts training programs, sponsors High Adventure awards, publishes specialized literature such as Hike Aids and The Trail Head and promotes participation in summer camp, in High Adventure activities such as backpacking, peak climbing, and conservation, and in other Council programs. Anyone who is interested in the GLAAC-HAT and its many activities is encouraged to direct an inquiry to the GLAAC-Camping Services or visit our web site at http://www.glaac-hat.org/. The GLAAC-HAT meets on the evening of the first Tuesday of each month at 7:30 pm in the Cushman Watt Scout Center, 2333 Scout Way, Los Angeles, CA 90026. These meetings are open to all Scouters. REVISIONS Jan 2016 General revision. Peak Bagger Peak list: Tom Thorpe removed Mt. San Antonio, added Blackrock Dick Rose Mountain. Mini-Peak Bagger list: removed Dawson Peak and Pine Mountain No. 1. Renamed "Suicide Peak" to "Suicide Rock". Updated "General Requirements" section. Jan 2005 New document incorporating Program Announcements 2 and 3. Prepared by Lyle Whited and composed by John Hainey. (Mt. Markham, summit trail) Peak Bagging Program and Awards -

The Mission Mountains Tribal Wilderness
University of Montana ScholarWorks at University of Montana Graduate Student Theses, Dissertations, & Professional Papers Graduate School 1995 A Confluence of sovereignty and conformity : the Mission Mountains Tribal Wilderness Diane L. Krahe The University of Montana Follow this and additional works at: https://scholarworks.umt.edu/etd Let us know how access to this document benefits ou.y Recommended Citation Krahe, Diane L., "A Confluence of sovereignty and conformity : the Mission Mountains Tribal Wilderness" (1995). Graduate Student Theses, Dissertations, & Professional Papers. 8956. https://scholarworks.umt.edu/etd/8956 This Thesis is brought to you for free and open access by the Graduate School at ScholarWorks at University of Montana. It has been accepted for inclusion in Graduate Student Theses, Dissertations, & Professional Papers by an authorized administrator of ScholarWorks at University of Montana. For more information, please contact [email protected]. A CONFLUENCE OF SOVEREIGNTY AND CONFORMITY: THE MISSION MOUNTAINS TRIBAL WILDERNESS by Diane L Krahe BA Bridgewater College, 1986 presented in partial fulfillment of the requirements for the degree of Master of Science The University of Montana 1995 Approved by; Chairperson Dean, Graduate School Date UMI Number: EP39757 All rights reserved INFORMATION TO ALL USERS The quality of this reproduction is dependent upon the quality of the copy submitted. In the unlikely event that the author did not send a complete manuscript and there are missing pages, these will be noted. Also, if material had to be removed, a note will indicate the deletion. UMT OiSiMflAtion Pubiishrig UMI EP39757 Published by ProQuest LLC (2013). Copyright in the Dissertation held by the Author. -

Mountain Ear
Mountain Ear THE MONTHLY NEWSLETTER OF THE ROCKY MOUNTAINEERS SEPTEMBER 1999 - The Rocky Mountaineers are dedicated to the exploration of ,Western Montana's mountains and other high peaks throughout the world* . * . ' - MEETING ' -- - Club Meetings: The Rocky Mountaineers normally meet the second wednesday of each month at the Missoula Public Library (lower level meeting room). The club does not meet during the summer months so that we can enjoy the long summer days &d explore our beloved mounta& - . - - - Our next meeting will be held on September 22 at 7 p.m. Please note that this is the fourth Wednesday in September. At the request of several members, the meeting was moved back to accommodate extended climbing trips over the Labor Day holiday. This meeting will be our annual "potluck" slide show, where we all show each other slides of our mountaineering exploits in the past year. A SPECIAL NOTE TO ALL MEMBERS: We are looking for new faces and energetic people to lead trips of all skill levels! Please contact Matt Grandy if you are interested in being a trip leader. TRIP CALENDER September 24-26, Fri-Sun, Three 12,000 footers and One 11,000+: Come climb with us in the Beartooth Mountains near Red Lodge. We'll ascend to Sundance Pass and set up a base camp. From here, we will scramble to Silver Run Peak (12,542 feet) and Point 12,500 (12,500 feet). The next day, we will do Mount Lockhart (1 1,647 feet) and Whitetail Peak (12,548 feet). When the Beartooth Mountains are viewed fiom the east, Whitetail Peak appears to be the highest peak in the range, and is often mistaken for Granite Peak - the highest peak in Montana. -

Montana Botany Notes Containing Description of New Species, List of Plants Not Heretofore Recorded from the State, and Notes on Disputed Species, 1910
University of Montana ScholarWorks at University of Montana University of Montana Bulletin: Biological Series, 1901-1910 Flathead Lake Biological Station 3-1-1910 Montana Botany Notes Containing Description of New Species, List of Plants Not Heretofore Recorded from the State, and Notes on Disputed Species, 1910 University of Montana (Missoula, Mont. : 1893-1913). Biological Station, Flathead Lake Marcus E. Jones Follow this and additional works at: https://scholarworks.umt.edu/umbiologicalseries Let us know how access to this document benefits ou.y Recommended Citation University of Montana (Missoula, Mont. : 1893-1913). Biological Station, Flathead Lake and Jones, Marcus E., "Montana Botany Notes Containing Description of New Species, List of Plants Not Heretofore Recorded from the State, and Notes on Disputed Species, 1910" (1910). University of Montana Bulletin: Biological Series, 1901-1910. 15. https://scholarworks.umt.edu/umbiologicalseries/15 This Book is brought to you for free and open access by the Flathead Lake Biological Station at ScholarWorks at University of Montana. It has been accepted for inclusion in University of Montana Bulletin: Biological Series, 1901-1910 by an authorized administrator of ScholarWorks at University of Montana. For more information, please contact [email protected]. BULLETIN. UNIVERSITY OF MONTANA Number 6 b BIOLOGICAL SERIES No. 15 MONTANA BOTANY NOTES CONTAINING Description of New Species, List of Plants Not Heretofore Recorded From the State, and Notes on Disputed Species, With Five Plates BY MARCUS E. JONES, A. M. Prepared From Material Collected at the University of Montana Biological Station UNIVERSITY OF MONTANA Missoula, Montana, U. S. A. March, 1910 Entered August 24, 1901, at Missonla, Montana, as second class matter, under act of Congress, July 16, 1894 BULLETIN, UNIVERSITY OF MONTANA Number 61 BIOLOGICAL SERIES No. -

The Armyworm and the Army Cutworm
NDSU EXTENSION E830 (Revised) The Armyworm and the Army Cutworm Revised by: Janet J. Knodel, Professor and Extension Entomologist rmyworm and army cutworm Travis J. Prochaska, Area Extension Crop Protection Specialist A feed on a wide variety of crops in North Dakota. Although the names are similar, these two insects are distinct, feeding at different times during the Armyworm growing season. Identifying and finding Mythimna unipuncta (Haworth) these insects, and recognizing when Lepidoptera: Noctuidae they become an economic threat will Description aid in successful pest management. The adult armyworm is a light brownish gray moth or “miller” (Figure 1) with a conspicuous white spot about the size of a pinhead on each front wing. When expanded, the wings are about 1½ inches across. Armyworm larvae (Figure 2) have five pairs of prolegs. Their color varies from pale green to tan in the early growth stage to dark green to black in later stages. The head capsule is brown with netlike patterns. Full-grown larvae are smooth, striped and almost hairless. They grow to a length of 1½ to 2 inches. A series of longitudinal stripes on the body are arranged as follows: n Thin, white, broken line down the middle of the black n Wide, dark, mottled stripe halfway down the side n Pale orange stripe with white border n Brownish mottled stripe n Another brownish mottled stripe slightly above the legs Life Cycle The armyworm does not survive North Dakota winters. Armyworm infestations Figure 1. Armyworm moth. (G. Fauske, NDSU) are due to moth migrations from the South. Heavy infestations in southern states produce large moth numbers that fly or are blown northward on southerly winds. -

Grizzly Bear Use of Army Cutworm Moths in The
GRIZZLYBEAR USE OF ARMYCUTWORM MOTHSIN THE YELLOWSTONEECOSYSTEM STEVENP. FRENCH,Yellowstone GrizzlyFoundation, 581 ExpeditionDrive, Evanston, WY 82930 MARILYNNG. FRENCH,Yellowstone GrizzlyFoundation, 581 ExpeditionDrive, Evanston, WY 82930 RICHARDR. KNIGHT,Interagency Grizzly Bear Study Team, ForestrySciences Laboratory,Montana State University, Bozeman, MT 59717 Abstract: The ecology of alpine aggregationsof army cutwormmoths (Euxoa auxiliaris) and the feeding behavior of grizzly bears (Ursus arctos horribilis) at these areas were studied in the Yellowstone ecosystem from 1988 to 1991. Army cutwormmoths migrateto mountainregions each summerto feed at nighton the nectarof alpineand subalpineflowers, and during the day they seek shelterunder various rock formations. Grizzly bears were observed feeding almost exclusively on moths up to 3 months each summerat the 10 moth-aggregationareas we identified. Fifty-one different grizzly bears were observed feeding at 4 of these areas during a single day in August 1991. Army cutworm moths are a preferred source of nutrition for many grizzly bears in the Yellowstone ecosystem and represent a high quality food that is available during hyperphagia. Int. Conf. Bear Res. and Manage.9(1):389-399 In 1986 the InteragencyGrizzly Bear StudyTeam bear RecoveryZone for the Yellowstoneecosystem. (IGBST)first observed radio-instrumented grizzly bears Additionalsurvey work occurred adjacent to the excavatingand presumably feeding in alpinetalus fields primarystudy area, in the GallatinNational Forest to located