Boom Clay (Rupelian, Oligocene) in Belgium by Tom VERHEYDEN
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Overzicht-Wijzigingen-Antwerpen.Pdf
Regio Antwerpen ........................................................................................................................3 Trams ......................................................................................................................................3 Lijn 2 Hoboken - Linkeroever ..............................................................................................3 Lijn 4 Hoboken – Sint Pietersvliet........................................................................................3 Lijn 8 Silsburg – Bolivarplaats .............................................................................................4 Lijn 9 Linkeroever - Eksterlaar .............................................................................................4 Lijn 11 Eksterlaar - Melkmarkt .............................................................................................5 Lijn 12 Sportpaleis – Bolivarplaats ......................................................................................5 Lijn 15 Mortsel – Linkeroever ..............................................................................................6 Bussen ....................................................................................................................................8 Lijn 9 Fruithoflaan – Rijnkaai ...............................................................................................8 Lijn 14 Vremde – Mortsel – Rooseveltplaats .......................................................................8 Lijn 19 Wenigerstraat -

Inventarisatie En Karakterisatie Van Verhoogde Concentraties Aan Natuurlijke Radionucliden Van Industriële Oorsprong in Vlaanderen
Aanvulling op de inventarisatie en karakterisatie van verhoogde concentraties aan natuurlijke radionucliden van industriële oorsprong in Vlaanderen Studie in opdracht van de Vlaamse Milieumaatschappij Johan Paridaens, Hans Vanmarcke SCK•CEN Departement Stralingsbeschermingsonderzoek BLG 916 Mol, Juni 2002 INHOUDSTAFEL SAMENVATTING.....................................................................................................................................1 Schematisch overzicht van de impact van de fostaatindustrie in Vlaanderen. .....................................2 Inleiding................................................................................................................................................3 Algemene werkwijze .............................................................................................................................3 I. HISTORISCHE FERRONIOBIUMPRODUCTIE, GENTSE KANAALZONE ....................................................4 I.1. Radioactiviteit in het productieproces ......................................................................................4 I.2. Terrein met ferroniobium slakken.............................................................................................4 I.2.1. Beschrijving, metingen................................................................................................................4 I.2.2. Bestemming, radiologische impact, aanbevelingen.....................................................................6 II. RADIUMHOUDENDE GIPSSTORTEN IN DE -

Gemeente Hemiksem RUP HET DORP Toelichtingsnota
Gemeente Hemiksem RUP HET DORP Toelichtingsnota Gemeente Hemiksem Sint-Bernardusabdij 1 2620 HEMIKSEM Grontmij Belgium Mechelen, maart 2012 247324_03_ontwerp, Revisie_c Verantwoording Titel : Gemeente Hemiksem RUP HET DORP Subtitel : Toelichtingsnota Projectnummer : 247324 Referentienummer : 247324_03_ontwerp Revisie : _c Datum : 6 maart 2012 Auteur(s) : Jeroen Bastiaens E-mail adres : [email protected] Gecontroleerd door : Jeroen Bastiaens Paraaf gecontroleerd : Goedgekeurd door : Paraaf goedgekeurd : Contact : Grontmij Belgium NV Stationsstraat 51 B-2800 Mechelen T +32 15 45 13 00 F +32 15 45 13 10 [email protected] www.grontmij.be 247324_03_ontwerp, Revisie_c Pagina 2 van 35 Gezien en definitief vastgesteld door de gemeenteraad in de vergadering van / / Op bevel, De Secretaris De Voorzitter Luc Schroyens Eddy de Herdt Zegel van de gemeente Verantwoordelijk Ruimtelijk Planner Jeroen Bastiaens 247324_03_ontwerp, Revisie_c Pagina 3 van 35 Inhoudsopgave LIJST VAN FIGUREN................................................................................................................... 6 1 INLEIDING.................................................................................................................... 7 1.1 Aanleiding tot de opmaak van het RUP........................................................................ 7 1.2 Afbakening van het plangebied .................................................................................... 7 2 RUIMTELIJKE CONTEXT........................................................................................... -

A New Classification of the Xanthoidea Sensu Lato
Contributions to Zoology, 75 (1/2) 23-73 (2006) A new classifi cation of the Xanthoidea sensu lato (Crustacea: Decapoda: Brachyura) based on phylogenetic analysis and traditional systematics and evaluation of all fossil Xanthoidea sensu lato Hiroaki Karasawa1, Carrie E. Schweitzer2 1Mizunami Fossil Museum, Yamanouchi, Akeyo, Mizunami, Gifu 509-6132, Japan, e-mail: GHA06103@nifty. com; 2Department of Geology, Kent State University Stark Campus, 6000 Frank Ave. NW, North Canton, Ohio 44720, USA, e-mail: [email protected] Key words: Crustacea, Decapoda, Brachyura, Xanthoidea, Portunidae, systematics, phylogeny Abstract Family Pilumnidae ............................................................. 47 Family Pseudorhombilidae ............................................... 49 A phylogenetic analysis was conducted including representatives Family Trapeziidae ............................................................. 49 from all recognized extant and extinct families of the Xanthoidea Family Xanthidae ............................................................... 50 sensu lato, resulting in one new family, Hypothalassiidae. Four Superfamily Xanthoidea incertae sedis ............................... 50 xanthoid families are elevated to superfamily status, resulting in Superfamily Eriphioidea ......................................................... 51 Carpilioidea, Pilumnoidoidea, Eriphioidea, Progeryonoidea, and Family Platyxanthidae ....................................................... 52 Goneplacoidea, and numerous subfamilies are elevated -

N-VA Borsbeek Heeft Nieuw Bestuur
Uw N-VA-schepenen (p. 2) BORSBEEK [email protected] I www.n-va.be/borsbeek I april 2019 I nr. 1 VERKIEZINGSDRUKWERK - V.U.: ANITA VAN DEN BULCK, WENIGERSTRAAT 61, 2150 BORSBEEK N-VA Borsbeek heeft nieuw bestuur Eind februari kozen de N-VA-leden van Borsbeek een nieuw afdelingsbestuur voor de komende drie jaar. Anita Van den Bulck werd verkozen als nieuwe voorzitter, Peggy Maebe als ondervoorzitter. Anita woont al bijna haar hele leven in Borsbeek en heeft al Sterke ploeg heel wat jaren op de politieke teller staan, eerst bij de Volksunie “Een voorzitter is niets waard zonder een stevige ploeg achter en nu bij de N-VA. De voorbije jaren was ze voorzitter van de zich”, aldus Anita. “Vandaar dat ik bijzonder trots ben op de gemeenteraad, nu is ze gemeenteraadslid. “Ik ben heel blij dat ik bestuursleden die met mij verkozen zijn. Dit bestuur is een de komende jaren onze dynamische, enthousiaste ploeg mag lei- gezonde mix van mensen met de nodige bestuurservaring en den”, zegt Anita. “De voorbije jaren mocht ik heel wat ervaring nieuwkomers die ongetwijfeld frisse en vernieuwende ideeën opdoen als gemeenteraadsvoorzitter en als Borsbekenaar in hart zullen aanbrengen. Samen willen we ervoor zorgen dat het beleid en nieren kan ik de noden van onze gemeente en haar inwoners van N-VA Borsbeek binnen onze gemeente nog verder wordt goed inschatten.” uitgedragen en erop toezien dat N-VA Borsbeek haar accenten kan leggen in het beleid van onze gemeente.” Peggy Maebe zal Anita bijstaan als ondervoorzitter. Zij versterkte drie jaar geleden het bestuur van N-VA Borsbeek. -

Overzicht Buitengewone Algemene Vergadering 16.12.2020 - 19 U - Digitaal
overzicht Buitengewone algemene vergadering 16.12.2020 - 19 u - digitaal Aanwezig 1. Deelnemers A. Gemeenten Aartselaar Bart Lambrecht Malle Dries De Vry Antwerpen Koen Laenens Mortsel Vera De Hert Antwerpen Peter Wouters Niel Kelly Breugelmans Boechout Kris Swaegers Ranst Luc Redig Boom Yani Brems Rumst Marc Verschueren Borsbeek Michael Bryon Schelle Pieter Smits Borsbeek Dis Van Berckelaer Schilde Laurenz Van Ginneken Brasschaat Joris Van Cauwelaert Schilde Philippe Verschueren Brecht Tatiana Vandekeere Schoten Sandra Lauryssen Edegem Joost Goris Stabroek Danielle Logiest Essen Dirk Konings Wijnegem Maarten Janssens Essen Gaston Van Tichelt Wuustwezel Jan Cools Hove Gerda Lambrecht Wuustwezel Sus Vissers Hove Bart Van Couwenberghe Zandhoven Paula Henderickx Kalmthout Wies Peeters Zoersel Bert Claessens Kontich Sophie Baeckelmans Zoersel Marcel De Vos Lint Harry Debrabandere Zwijndrecht Mariam Amal Zwijndrecht Greet Bergmans verontschuldigd: Hemiksem Jill Van Wijnsberghe Kapellen Nancy Van Huffelen Wommelgem Lode Stevens B. OCMW’s, intergemeentelijke verenigingen en anderen Aartselaar Guido D’eer Stabroek Danielle Logiest Boechout Kris Swaegers Stabroek Jan Schouwaerts Borsbeek Michael Bryon Wijnegem Maarten Janssens Borsbeek Dis Van Berckelaer Wuustwezel Koen Van Putte Brasschaat Joris Van Cauwelaert Zandhoven Paula Henderickx Brecht Tatiana Vandekeere Zoersel Bert Claessens Edegem Joost Goris Zoersel Marcel De Vos Hove Gerda Lambrecht Zwijndrecht Mariam Amal Hove Bart Van Couwenberghe Zwijndrecht Greet Bergmans Kalmthout Wies Peeters -

Introduction Day To
Amsterdam - Brussels - 8 dagen DUTCH BIKETOURS - EMAIL: [email protected] - TELEPHONE +31 (0)24 3244712 - WWW.DUTCH-BIKETOURS.COM Amsterdam - Brussels 8 days, € 759 Introduction This international bicycle holiday starts in the heart of the Dutch capital and finishes in the epicenter of Europe, Brussels. It’s a one-way route, 340 kilometres long, past a string of exceptional and historically precious Dutch and Flemish towns. In- between towns, enjoy the peace and quiet of the Dutch and Flemish countryside with age-old farmhouses and dominating church spires. You will follow a well- signposted route that is as good as flat, low traffic or even car free. Only towards the very end does it get slightly hilly. The towns themselves are perfect for touring by bike, ensuring total freedom and mobility: no traffic jams and you can “park” anywhere. Day to Day Day 1 Arrival Amsterdam Your cycling holiday starts in Amsterdam, one of the crown-jewels of Europe. With so much to see and do, you couldn’t wish for a more vibrant way to kick off your holiday. If you arrive early, make sure to go for a walk to marvel at the townhouses in the 17th- century canal district (listed as World Heritage). Have you got time and energy left? Opt to saunter down the infamous red-light district which attracts millions of curious tourists every year; or enjoy the street performers on friendly Rembrandtplein. Day 2 Amsterdam – Gouda 79 km The first leg of the route takes you through the Green Heart of Holland, abounding in water. -
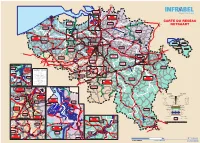
Carte Du Reseau Netkaart
AMSTERDAM ROTTERDAM ROTTERDAM ROOSENDAAL Essen 4 ESSEN Hoogstraten Baarle-Hertog I-AM.A22 12 ANTWERPEN Ravels -OOST Wildert Kalmthout KALMTHOUT Wuustwezel Kijkuit Merksplas NOORDERKEMPEN Rijkevorsel HEIDE Zweedse I-AM.A21 ANTW. Kapellen Kaai KNOKKE AREA Turnhout Zeebrugge-Strand 51A/1 202 Duinbergen -NOORD Arendonk ZEEBRUGGE-VORMING HEIST 12 TURNHOUT ZEEBRUGGE-DORP TERNEUZEN Brasschaat Brecht North-East BLANKENBERGE 51A 51B Knokke-Heist KAPELLEN Zwankendamme Oud-Turnhout Blankenberge Lissewege Vosselaar 51 202B Beerse EINDHOVEN Y. Ter Doest Y. Eivoorde Y.. Pelikaan Sint-Laureins Retie Y. Blauwe Toren 4 Malle Hamont-Achel Y. Dudzele 29 De Haan Schoten Schilde Zoersel CARTE DU RESEAU Zuienkerke Hamont Y. Blauwe Toren Damme VENLO Bredene I-AM.A32 Lille Kasterlee Dessel Lommel-Maatheide Neerpelt 19 Tielen Budel WEERT 51 GENT- Wijnegem I-AM.A23 Overpelt OOSTENDE 50F 202A 273 Lommel SAS-VAN-GENT Sint-Gillis-Waas MECHELEN NEERPELT Brugge-Sint-Pieters ZEEHAVEN LOMMEL Overpelt ROERMOND Stekene Mol Oostende ANTWERPEN Zandhoven Vorselaar 50A Eeklo Zelzate 19 Overpelt- NETKAART Wommelgem Kaprijke Assenede ZELZATE Herentals MOL Bocholt BRUGGE Borsbeek Grobbendonk Y. Kruisberg BALEN- Werkplaatsen Oudenburg Jabbeke Wachtebeke Moerbeke Ranst 50A/5 Maldegem EEKLO HERENTALS kp. 40.620 WERKPLAATSEN Brugge kp. 7.740 Olen Gent Boechout Wolfstee 15 GEEL Y. Oostkamp Waarschoot SINT-NIKLAAS Bouwel Balen I-AM.A34 Boechout NIJLEN Y. Albertkanaal Kinrooi Middelkerke OOSTKAMP Evergem GENT-NOORD Sint-Niklaas 58 15 Kessel Olen Geel 15 Gistel Waarschoot 55 219 15 Balen BRUGGE 204 Belsele 59 Hove Hechtel-Eksel Bree Beernem Sinaai LIER Nijlen Herenthout Peer Nieuwpoort Y. Nazareth Ichtegem Zedelgem BEERNEM Knesselare Y. Lint ZEDELGEM Zomergem 207 Meerhout Schelle Aartselaar Lint Koksijde Oostkamp Waasmunster Temse TEMSE Schelle KONTICH-LINT Y. -

Downloaded from Brill.Com09/26/2021 12:42:37AM Via Free Access 102 A
Contributions to Zoology, 72 (2-3) 101-104 (2003) SPB Academic Publishing bv, The Hague Decapod crustaceans from the Oligocene of the Ligure Piemontese Basin, northern Italy Antonio De Angeli¹ & Sergio Marangon² 'Associazione Amici del Museo civico G. Zannato Piazza Marconi 15, 1-36075 Montecchio Maggiore 2 (Vicenza), Italy; c/o Via Anemone 8, 1-20090 Segrate (Milano), Italy Keywords: Crustacea, Decapoda, Oligocene, Italy M. Abstract species postulosus. Subsequently, new material housed in the collections of the Museo civico at Lithologically, the decapod-bearing levels of Rupelian (Oligo- Ovada and the Museo regionale di Scienze naturali cene) in the Ligure Piemontese Basin, are characterizedby age at Torino, was studied by Allasinaz (1987), who an alternation of greyish blue marls with nodular elements and described'Calappilia mainii Alassinaz, C. vicetina silt-rich, occasionally sandy, marls. The fossil studied for the and Portunus Fabiani, Calappa sp. monspeliensis present note have been collected mainly from pebblesor nodules Milne The additions in- which (A. Edwards). most recent were eroded out of the higher levels exposed of this clude rostratus & De sedimentary complex, referred to as the ‘Formazione di Mol- Cherpiocarcinus Marangon are’. These levels overli the ‘Formazioni continentali delle Brecce Angeli (Fig. 1/1) and Calappilia verrucosa A. Milne di studied al. Costa Cravara e Charrier et (1964) Pianfolco’, by Edwards (Fig. 1/2) from Cherpione (Pianfolco) (see and dated as Early Rupelian. The levels yielding brachyuran Marangon & De Angeli, 1997; De Angeli & Maran- fossils were attributed by Allasinaz (1987) to the transition bet- in Paralbunea gon, press) and galantensis from ween the ‘Formazione di Molare’ and the overlying ‘Marne di Contra Galanti (Cassinelle) (see De Angeli & Maran- Rigoroso’, and by Bianco (1985) and Balossino& Bianco (1986) to the biozone of the foraminifer Operculina complanata (De- gon, 2001). -

How to Reach Our Antwerp Offices? Prins Boudewijnlaan 41, 2650 Edegem Tel: 00 32 (0)3 281 44 88
How to reach our Antwerp offices? Prins Boudewijnlaan 41, 2650 Edegem Tel: 00 32 (0)3 281 44 88 By car Coming from Brussels-Mechelen E19 Continue on the E19 Brussel – Antwerpen. Take Exit 7- Kontich Take right at the end of the exit lane, direction Kontich/Antwerpen. Go straight ahead. You now arrive on the Prins Boudewijnlaan. You reach the Quintessence Antwerp office at nr 41 approximately 1,2 km past the second traffic lights. Enter the parking lot by turning right and park freely on the reserved Quintessence parking. Coming from Brussels A12 Continue on the A12 Brussel/Antwerpen. Watch out for speed control! Just after the Rupeltunnel: take the Exit Boom/Aartselaar/Kontich. Keep on the right lane. You are now on the Boomsesteenweg. At the first traffic light, turn right in the direction of Aartselaar. Go straight ahead. Turn right at the T-crossroad. Continue. Pass the traffic lights in the direction of Kontich- Rumst. At approx. 1 km turn left at the express way N171. (direction E19Antwerpen/Brussel). Go straight a head. (pass E19) You now arrive on the Prins Boudewijnlaan. You reach the Quintessence Antwerp office at nr 41 approximately 1,2 km past the second traffic lights. Enter the parking lot by turning right and park freely on the reserved Quintessence parking. Quintessence Consulting NV Prins Boudewijnlaan 41 2650 Edegem Tel: 03/281 44 88 Fax: 03/281 35 57 Boulevard Général Wahis 3 1030 Bruxelles Tel: 02/705 28 48 Fax: 02/705 28 50 [email protected] www.quintessence.be Coming from Ghent-Bruges-Kortrijk E17/ Knokke-Zelzate E34 After the Kennedy tunnel follow the signs “E19 Brussel-Mechelen” Pass the Craeybeckxtunnel and continue on the E19 Brussel. -

Kontich (ANT) ADEDE Archeologisch Rapport 150
2017 ARCHEOLOGIENOTA Antwerpsesteenweg – Kontich (ANT) ADEDE Archeologisch Rapport 150 JANSSENS D. VAN HUFFEL C. ADEDE ARCHEOLOGISCH RAPPORT 150 Archeologienota Antwerpsesteenweg te Kontich (Antwerpen). JANSSENS DAVID & VAN HUFFEL CEDRIC Colofon Uitgever ADEDE bvba Jaar van uitgave 2017 Plaats van uitgave Gent Redactie Bart De Smaele, Hadewijch Pieters ISSN 2033-6810 Niets uit deze uitgave mag worden verveelvoudigd en/of openbaar gemaakt door middel van druk, fotokopie, microfilm of op welke andere wijze dan ook, zonder voorafgaande schriftelijke toestemming van ADEDE bvba. Inhoudsopgave 1 Administratieve fiche ................................................................................................................... - 5 - 2 Bureauonderzoek ....................................................................................................................... - 11 - 2.1 Archeologische voorkennis ............................................................................................... - 11 - 2.2 Aanleiding van het onderzoek ........................................................................................... - 11 - 2.3 Doel van het onderzoek .................................................................................................... - 11 - 2.4 Huidige situatie projectgebied .......................................................................................... - 12 - 2.5 Beschrijving geplande werken ........................................................................................... - 15 - 2.6 Randvoorwaarden -
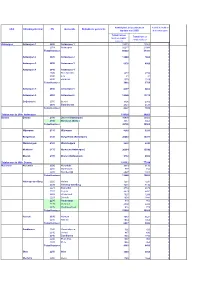
Update Mei 2005) Brievenbussen
Aantal part. brievenbussen Aantal betrokken UKA Uitreikingskantoor PN Gemeente Betrokkene gemeente (update mei 2005) brievenbussen Totaal bussen Totaal bussen (met en zonder zonder sticker sticker) Antwerpen Antwerpen 1 2000 Antwerpen 1 28375 19365 0 2018 Antwerpen 32217 21938 0 Totaal kantoor: 60592 41303 0 0 Antwerpen 2 2020 Antwerpen 2 12580 7860 0 0 Antwerpen 3 2030 Antwerpen 3 6178 4959 0 0 Antwerpen 4 2040 Antwerpen 4 0 2040 Berendrecht 2873 2706 0 2040 Lillo 31 31 0 2040 Zandvliet 1078 1019 0 Totaal kantoor: 3982 3756 0 0 Antwerpen 5 2050 Antwerpen 5 8317 6862 0 0 Antwerpen 6 2060 Antwerpen 6 19248 13176 0 0 Zwijndrecht 2070 Burcht 3406 2910 0 2070 Zwijndrecht 4821 4125 0 Totaal kantoor: 8227 7035 0 0 Totalen van de UKA Antwerpen 119124 84951 0 Deurne Deurne 2100 Deurne (Antwerpen) 35909 26502 0 2150 Borsbeek (Antw.) 4887 4063 0 Totaal kantoor : 40796 30565 0 0 Wijnegem 2110 Wijnegem 4218 3231 0 0 Borgerhout 2140 Borgerhout (Antwerpen) 20810 15977 0 0 Wommelgem 2160 Wommelgem 5422 4293 0 0 Merksem 2170 Merksem (Antwerpen) 20314 15546 0 0 Ekeren 2180 Ekeren (Antwerpen) 9752 8302 0 0 Totalen van de UKA Deurne 101312 77914 0 Herentals Herentals 2200 Herentals 9884 8136 0 2200 Morkhoven 777 748 0 2200 Noorderwijk 2027 1918 0 Totaal kantoor: 12688 10802 0 0 Heist-op-den-Berg 2220 Hallaar 1281 1231 0 2220 Heist-op-den-Berg 7401 7192 0 2221 Booischot 2734 2679 0 2222 Itegem 2279 2201 0 2222 Wiekevorst 1327 1299 0 2223 Schriek 1902 1842 0 2235 Houtvenne 818 766 0 2235 Hulshout 2392 2222 0 2235 Westmeerbeek 916 773 0 Totaal kantoor: 21050