Benifallet, Tarragona
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

BIM 3Er Quadrimestre 2017
BUTLLETÍ D’INFORMACIÓ Núm. 8 MUNICIPAL 3a Època 3R Quadrimestre AJUNTAMENT DE 2017 MÓRA LA NOVA MÓRA LA NOVA MUNICIPAL Móra la Nova aprova un Les festes de Nadal una cita pressupost consensuat de més plena d’activitats per a tothom de 3,5M d’euros Una Sant Silvestre aquàtica i el Més visitants a la 186a Fira, que Club de Twirling moranovenc en ha presentat moltes novetats molta projecció Pavelló Firal 1 d’Octubre, el nou Commemoració del Dia nom del recinte després dels internacional per a l’eliminació atacs policials de la violència de gènere MÓRA LA NOVA MUNICIPAL Editorial Número 8 Salutació Des de finals d’octubre, ERC governa 3a Època en minoria a Móra la Nova. Les fortes 3r Quadrimestre 2017 tensions nacionals i els diferents punts de vista sobre el procés van fer insostenible el pacte de govern. Però Edita el trencament ha estat d’una forma Ajuntament de Móra la Nova civilitzada i sensata, com haurien de ser totes les relacions que finalitzen. Coordinació Hem de conviure en el mateix poble sense animadversió. Eva Vallespí El governar en minoria ens ha Francesc X. Moliné obligat a negociar i a pactar uns pressupostos pel 2018 amb l’oposició. Redacció Fruit d’aquesta negociació, s’han Roger Font i Montornés aprovat amb els vots favorables del PDeCat i del PSC després de recollir algunes de les seves esmenes. www.riberaproduccions.com D’altra banda, al mes d’octubre va tenir lloc la 186 Fira de Móra la Nova. Hem introduït alguns canvis fruit de la pressió urbanística que ens Col·laboradors han servit per reorganitzar la Fira i per traure’n una part de l’àmbit Antonio Veciana (meteorologia) urbà, minimitzant molèsties. -

El Republicanisme a Les Terres De L Lebre En Temps
EL REPUBLICANISME A LES TERRES DE L’EBRE EN TEMPS DEL CNR EL REPUBLICANISME A LES TERRES DE L’EBRE EN TEMPS DEL CNR JOSEP SÁNCHEZ CERVELLÓ 341 Recerca, 14 (2012), p. 339-373. ISSN 1135-6014 EL REPUBLICANISME A LES TERRES DE L’EBRE EN TEMPS DEL CNR RESUM A les Terres de l’Ebre, fins 1907, el republicanisme fou dirigit per la Unión Republicana que tingué més lligams amb el nord de la província de Castelló i València que amb Barcelona, visualitzant-se, a més, una estreta relació amb el ‘blasquisme’. A partir del sorgiment de Solidaritat Catalana, el republicanisme a l’Ebre tingué dos eixos fonamentals, l’un al Baix Ebre/Montsià i l’altre al districte de Gandesa que incloïa la Ribera d’Ebre, la Terra Alta i la part del Priorat compresa dins la Diòcesi de Tortosa. A les dues comarques del sud es compatibilitzà el blasquisme i la influència de Solidaritat Catalana en una síntesi que donà lloc al marcel·linisme; mentre que a les comarques del nord l’elecció del diputat Joan Caballè Goyeneche, l’any 1907 per Solidaritat Catalana i vinculat a la Unió Federal Nacionalista Republicana, portà el districte de Gandesa a una vinculació més estreta amb el republicanisme catalanista representat pel Foment Republicà de Reus, tot i que també en aquesta zona el marcel·linisme tingué un fort arrelament. De fet el marcel·linisme fou el corrent polític d’esquerres majoritari a l’Ebre fins la Guerra Civil. Paraules clau: republicanisme, Marcel·lí Domingo, Joan Caballé, Terres de l’Ebre RESUMEN El republicanismo en las Terres de l’Ebre estuvo liderado, hasta 1907 por Unión Republicana, partido de mayor relación con el norte de la provincia de Castellón y Valencia que con Barcelona, y que mostraba una estrecha relación con el ‘blasquismo’. -

Desplegament Fibra Òptica 2019-2021 Demarcació De Tarragona
Desplegament 2020-2022 demarcació de Tarragona Cristina Campillo i Cruellas – Gencat.cat Jaume Vidal González – Diputació de Tarragona Versió 1 – Gener de 2021 Desplegament 2020 2 Desplegament 2020 (I). Capitals de comarca. El 2020, s’ha fet el desplegament de capitals de comarca, obres promogudes per la Secretaria de Polítiques Digitals (SPD). Llegenda: Xarxa ja existent (cable propi) Xarxa ja existent (disponibilitat de fibres a cable de tercers) Xarxa desplegada per la SPD el 2020 Xarxa desplegada per la XOC el 2020 Calendari de recepció d’obres: • El Vendrell – Valls: 31/12/2020. • Valls - Montblanc: 31/12/2020. • Tortosa – Gandesa: 31/12/2020. • Mora la Nova – Falset: 31/01/2021. A disposició del mercat majorista gener/2021 (22/gener) 3 Desplegament 2020 (II). Instruments de comercialització. Llegenda: Xarxa ja existent (cable propi) Xarxa ja existent (disponibilitat de fibres a cable de tercers) Xarxa desplegada per la SPD el 2020 Xarxa desplegada per la XOC el 2020 Instruments de comercialització: Xarxa desplegada per la SPD el 2020 • Preu públic CTTI de lloguer de conductes: 0,53 €/m/any amb bonificacions de fins el 50% en funció de la densitat i número d’habitants del terme municipal. • Nou preu públic CTTI de lloguer de fibres fosques (finals gener) Sol·licituds via el Punt d’Informació Únic (PIU) • https://politiquesdigitals.gencat.cat/ca/tic/piu/ Xarxa desplegada per la XOC el 2020 • Oferta majorista de lloguer de fibres fosques • Oferta majorista de serveis actius • https://www.xarxaoberta.cat/ 4 Desplegament 2020 (II) Els -
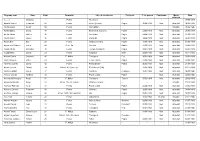
Cognom, Nom Nom Edat Cementiri Lloc De Residència Professió C. De
Cognom, nom Nom Edat Cementiri Lloc de residència Professió C. de guerra Condemna Motiu Data defunció (..., ...), Domingo - Fossa No consta - - - - 03/05/1939 Abadia Gómez Francesc 64 Fossa Falset (Priorat) Pagès 10/04/1939 Mort Afusellat 06/05/1939 Abellà Costa Serafí 55 Fossa Móra d'Ebre - - - Malaltia 05/02/1940 Abella Mateu Josep 46 Fossa Barberà de la Conca Pagès 28/02/1939 Mort Afusellat 20/03/1939 Abellà Trillas Maties 36 Fossa Montblanc Pagès 24/04/1939 Mort Afusellat 31/05/1939 Abelló Cabré Jaume 36 Fossa Poboleda Pagès 14/04/1939 Mort Afusellat 06/05/1939 Adrian Bes Blai 28 Fossa Vilalba dels Arcs Pagès 11/05/1939 Mort Afusellat 15/06/1939 Agustench Solanes Josep 49 Secc. 5a Mont-ral Barber 12/05/1939 Mort Afusellat 08/08/1939 Aixalà Abelló Salvador 30 Fossa Torroja del Priorat Pagès 09/05/1939 Mort Afusellat 14/07/1939 Aixalà Masó Josep 29 Fossa Amposta Xofer 20/07/1939 Mort Afusellat 16/11/1939 Alabau Garcia Sebastià 45 P. Militar Tarragona Peó 16/02/1939 Mort Afusellat 28/02/1939 Alba Carbonell Rafel 23 Fossa Jesús i Maria Pagès 21/02/1940 Mort Afusellat 20/06/1941 Alberich Cubells Josep 42 Fossa Palma d'Ebre Paleta 06/06/1939 Mort Afusellat 21/07/1939 Albert Colomé Dionisi 29 Nínxol 40 - Secc. 6a El Catllar de Gaià - 11/08/1939 Mort Afusellat 15/11/1939 Albiñana Matas Josep 37 Fossa Cambrils Perruquer 16/01/1940 Mort Afusellat 04/09/1940 Alcover Llorens Ambrosi 32 Fossa Pla de Cabra Pagès - Mort Afusellat 24/08/1940 Alcoverro Berenguer Àngel 54 P. -

Consorci Per Al Desenvolupament Del Baix Ebre I Montsià)
Olive trees field. Mas de Barberans, Montsià (Consorci per al Desenvolupament del Baix Ebre i Montsià) Consorci per al Desenvolupament del Baix Ebre i Montsià Area: Baix Ebre: l’Aldea, Aldover, Alfara de Carles, l’Ametlla de Mar, l’Ampolla, Benifallet, Camarles, Deltebre, Paüls, el Perelló, Roquetes, Tivenys, Tortosa and Xerta. 1.738,1 km2 Montsià: Alcanar, Amposta, Freginals, la Galera, Godall, Mas de Barberans, Masdenver- Population: ge, Sant Carles de la Ràpita, Sant Jaume d’Enveja, Santa Bàrbara, la Sénia and Ulldecona. 150.250 inhabitants Population density: 86,44 inhab./km2 Number of municipalities: 26 municipalities The territory Contact details Consorci per al Desenvolupament del Baix Ebre i Montsià works along the most Southern regions with a privileged position as a junction between Catalonia, Valencia and Aragon; Office and between two dynamic communication axis: the Mediterranean axis and the river Ebre Corridor. Consorci per al Desenvolupament del Baix Ebre i Montsià In landscape terms, the region has two differentiated areas. The mountains range, where Els Ports, Montsià range and Cardó stand out; the inland and coastal plains; and finally the Pl. Alfons XII, 7. 1r B riverbank and the Delta; all of them configured by one of the most characteristic elements of 43500 Tortosa both regions: Ebre river, which has played a decisive role in the economic and human shaping Phone (+34) 977 44 21 16 of both regions. This landscape has been recently recognised as a Biosphere Reserve, together with Terra Alta and Ribera d’Ebre; and it is protected by different figures, the most relevant www.leader.cat ones are: Ebre Delta Natural Park and Els Ports Natural Park. -

Publicación De Una Solicitud De Registro Con Arreglo Al Apartado 2
29.1.2003ES Diario Oficial de las Comunidades Europeas C 22/5 Publicación de una solicitud de registro con arreglo al apartado 2 del artículo 6 del Reglamento (CEE) no 2081/92 del Consejo relativo a la protección de las indicaciones geográficas y de las denominaciones de origen (2003/C 22/03) Esta publicación otorga un derecho de oposición con arreglo al artículo 7 del citado Reglamento. Cualquier oposición a esta solicitud debe enviarse por mediación de la autoridad competente de un Estado miembro en un plazo de seis meses a partir de la presente publicación. La publicación está motivada por los elementos que se enumeran a continuación, principalmente el punto 4.6, según los cuales se considera que la solicitud está justificada en virtud del Reglamento (CEE) no 2081/92. REGLAMENTO (CEE) No 2081/92 DEL CONSEJO SOLICITUD DE REGISTRO: ARTÍCULO 5 DOP ( ) IGP (x) Número nacional del expediente: 75 1. Servicio competente del Estado miembro Nombre: Subdirección General de Denominaciones de Calidad y Relaciones Interprofesionales y Contractuales Dirección General de Alimentación Subsecretaría de Agricultura Ministerio de Agricultura, Pesca y Alimentación. Dirección: Paseo Infanta Isabel, 1, E-28071 Madrid Tel. (34-91) 347 53 97 Fax (34-91) 347 54 10. 2. Agrupación solicitante 2.1. Nombre: a) Agrofruit Export, SA b) Cooperativa Exportadora d'Agris d'Alcanar c) Cooperativa de Bítem d) El Progrés, SCCL. 2.2. Dirección: a) Polígon Industrial Baix Ebre, Carré A, E-43500 Tortosa b) Carretera Nova 77-87, E-43530 Alcanar c) Colònia Gàsol, 132, E-43510 Bítem d) Av. Lluís Companys s/n, E-43512 Benifallet. -

Segregacions: Un Problema De Fronteres
SEGREGACIONS: UN PROBLEMA DE FRONTERES ESTUDI HISTÒRICO-JURÍDIC A TRAVÉS DEL PERELLÓ I LES TERRES DE L’EBRE El Perelló 2014 1 2 ÍNDEX Relació d'abreviatures pàg. 5 1. Introducció pàg. 7 2. Origen i evolució del municipi pàg.13 2.1 Teories pàg.13 2.2 Evolució dels municipis a les Terres de l'Ebre pàg.14 3. El municipi liberal pàg.19 4. Segregacions durant el segle XIX pàg.21 4.1 Marc legal pàg.21 4.2 Enumeració i descripció essencial de les segregacions a les pàg.27 Terres de l'Ebre 4.3 Anàlisi concret de la segregació de l'Ametlla de Mar pàg.28 respecte del Perelló 5. Segregacions durant el segle XX pàg.37 5.1 Marc legal pàg.37 5.2 Enumeració i descripció essencial de les segregacions a les pàg.42 Terres de l'Ebre 5.3Anàlisi concret de la segregació de l'Ampolla respecte del Perelló pàg.45 6. Comparativa entre segregacions del s. XIX respecte al s. XX pàg.61 Legislació aplicable Parts legitimades per instar la segregació Òrgan competent Requisits legals Participació dels actors en el procés Adequació a la realitat social 7. Consideracions finals pàg.67 8. Bibliografia pàg.69 9. Fonts documentals pàg.73 Annexos pàg.75 3 4 Relació d'abreviatures BOE Butlletí Oficial de l'Estat CE Constitució Espanyola DOGC Diari Oficial de la Generalitat de Catalunya FJ Fonament Jurídic Sr Senyor TC Tribuna Constitucional TS Tribunal Suprem Pàg. Pàgina Pàgs. Pàgines 5 6 1. INTRODUCCIÓ En l'actualitat ens trobem immersos en una greu crisi econòmica que ha provocat que es qüestioni l'organització i el model de la societat contemporània. -

S8GCI0N PROVINCIAL T LOCAL Ráo & Madrid Pasando Por Valencia, Donde Se Agre Ca Al Notario De Cambriis D
IMPUESTO DE CONSUMOS el'caciquismo, ha vencido, según se dice, el candi Según se nos ba notificado, el solemne septena tantes de entre sus compañeros que más se distin dato ministerial, gracias á los esfuerzos y á tos rio de los Dolores que debia empezar mañana en guen por su significación democrática. EN lA PROVINCIA. medios empleados para qae nacieran de las urnas la parroquial iglesia de San Juan Bautista, se tras Distribución de cupos de consumos y cereales qne papeletas con el nombre del Sr. Ballester. lada para después del novenario de San José, por Ha sido declarada la caducidad de las minas que por las especies comprendidas en la Tarifa 1,* Se asegura por aquí que en los pueblos peque no encontrarse bien de salud el Rdo. Padre Espe se expresan á continuación: del referido impuesto corresponde satisfacer á ños como Tamarit, Pobla y otros del distrito, apa ranza, quien tiene á su cargo los sermones del «Sultana» y «Virgen María», de cobre, sitas en los pueblos de la provincia en el ejercicio pró recen votando todos los electores al candidato sa- referido septenario. el término de Alforja, y concedidas á D. Eduardo ximo de 1882 á 83 y segundo semestre del ac gastino, aun los imposibilitados, y como ya esta Domínguez. tual, con arreglo á la ley é instrucción de 31 de mos acostumbrados á presenciar impasibles ciertas Pasan de ciento cincuenta los objetos que han «Rica plomiza» y «Mana», de plomo, término diciembre último, sin perjuicio de las alteracio cosas, de ahí que lleguen á ser moneda corriente regalado varias familias de esta capital para la de la Selva, y concedidas á D. -

FIESTA Y CULTURA Un Destacado Carácter Histórico De Diversidad Y Tolerancia
LA MÓRA MORISCA EN MÓRA D’EBRE: TRES VISIONES, UN SOLO MUNDO 7 y 8 de julio de 2012 Los vecinos del mercadal y de los barrios cristiano, judío y morisco ambientan las calles y vestidos de época, recrean un momento histórico de convivencia pacífi ca. Artesanos de ofi cios antiguos, espectáculos teatrales y de calle, desfi les, torneos, paradas y productos tradicionales, invitan a los visitantes a disfrutar de una fi esta con FIESTA Y CULTURA un destacado carácter histórico de diversidad y tolerancia. FIESTA DE SAN ANTONIO EN ASCÓ SEMANA SANTA EN FLIX: PROCESIÓN DEL ENCUENTRO Y LAS DESMAS Enero de 2013 Domingo de Pascua Declarada fi esta de interés comarcal y nacional Declarada fi esta de interés comarcal La Fiesta de San Antonio en Ascó ha mantenido vivas unas tradiciones que se pierden en Una apretada agenda de actos laicos, como las coplas o el baile de jotas en la ermita el tiempo y que la hacen singular: mantenimiento de la pliega, las Tres Vueltas, la distin- el Lunes de Pascua, y actos religiosos, entre los que destaca la Procesión del En- ción de las fi guras del clavario y los mayorales, las carreras de caballos, burros y machos, cuentro. La encabezan los Armados con la Samaritana seguidos por las banderas del encender la hoguera y bailar alrededor la jota de Ascó, con música y danza propias. Sagrado Corazón, la madre de dios del Remedio, San Antonio y el Santísimo. Cierra la Virgen, con la Cofradía de los Dolores y un grupo de esclavas. En la plaza Mayor se FUEGO Y EMPUJE; SAN ANTONIO EN LA TORRE DE L’ESPANYOL hace la representación del encuentro de la Virgen con Cristo resucitado y la Samari- Enero de 2013 tana recita Las Desmas, que acaban con la rotura del cántaro. -

El Cobre De Linares (Jaén) Como Elemento Vinculado Al Comercio Fenicio En El Calvari De El Molar (Tarragona) Ignacio Montero-Ruiz, Núria Rafel, M
JUNTA DE ANDALUCÍA. CONSEJERÍA DE CULTURA Y DEPORTE Conjunto Arqueológico Dólmenes de Antequera ISSN 2172-6175 Depósito Legal: SE 8812-2011 Menga es una publicación anual del Conjunto Arqueológico Dólmenes de Antequera (Consejería de Cultura y Deporte de la Junta de Andalucía). Su objetivo es la difusión internacional de trabajos de investigación científicos de calidad relativos a la Prehistoria de Andalucía. Menga se organiza en cuatro secciones: Dossier, Estudios, Recensiones y Crónica. La sección de Dossier aborda de forma monográfica un tema de investigación de actualidad. La segunda sección tiene un propósito más general y está integrada por trabajos de temática más heterogénea. La tercera sección incluye reseñas de libros y otros eventos (tales como exposiciones científicas, seminarios, congresos, etc.). La última sección denominada como Crónica recogerá las actuaciones realizadas por el Conjunto Arqueológico Dólmenes de Antequera en la anualidad anterior. Menga está abierta a trabajos inéditos y no presentados para publicación en otras revistas. Todos los manuscritos originales recibidos serán sometidos a un proceso de evaluación externa y anónima por pares como paso previo a su aceptación para publicación. Excepcionalmente, el Consejo Editorial podrá aceptar la publicación de traducciones al castellano y al inglés de trabajos ya publicados por causa de su interés y/o por la dificultad de acceso a sus contenidos. Menga is a yearly journal published by the Dolmens of Antequera Archaeological Site (the Andalusian Regional Government Ministry of Culture and Sport). Its aim is the international dissemination of quality scientific research into Andalusian Prehistory. Menga is organised into four sections: Dossier, Studies, Reviews and Chronicle. -

Reducció De L'atur I Migracions Recents a Les Terres De L'ebre
REDUCCIÓ DE L'ATUR I MIGRACIONS RECENTS A LES TERRES DE L'EBRE Autors: Juan Antonio Duro (CELIR i Departament d’Economia, URV) i Josep Maria Piñol (CELIR i FURV) 1. EVOLUCIÓ DEMOGRÀFICA: UNA NOVA FASE RECESSIVA A L'EBRE El present document intenta posar sobre la taula alguns elements d'anàlisi comparada en l'evolució demogràfica i de l'atur en els darrers anys a les Terres de l’Ebre. Més que un detall excessiu en algunes dades, es busca contrastar unes dinàmiques fonamentalment interanuals acumulades en aquests dos components i prenent la dades obertes disponibles. Els objectius són: a) Intentar identificar una possible correlació entre migració i atur registrat, incloent-hi les possibles especificitats locals. b) Confirmar si existeix una disminució de l'atur relacionades amb noves dinàmiques d'emigració i a quins perfils es vincula majoritàriament. Des del punt de vista demogràfic, les Terres de l'Ebre es mouen habitualment entre l'estancament i la regressió. Les dades recents indiquen que, després d'un ascens puntual, en els darrers anys —i paral·lelament a la major incidència de la crisi— la regió entra en una nova fase regressiva i desfavorable en relació amb Catalunya (taula 1). Taula 1. EVOLUCIÓ DEMOGRÀFICA RECENT Àmbit 2011 2012 2013 2014 2015 2016 Var % var. 2011-16 2011-16 Baix Ebre 82.634 83.125 81.514 80.637 79.748 78.977 -3.657 -4,43 Montsià 72.261 72.121 71.577 69.613 68.524 67.646 -4.615 -6,39 Ribera d'Ebre 23.889 23.867 23.477 22.925 22.723 22.471 -1.418 -5,94 Terra Alta 12.713 12.713 12.310 12.119 11.872 11.761 -952 -7,49 Terres de l'Ebre 191.631 191.826 188.878 185.294 182.867 180.855 -10.776 -5,62 Catalunya 7.539.618 7.570.908 7.553.650 7.518.903 7.508.106 7.522.596 -17.022 0,22 Font: elaboració pròpia a partir de dades de l'Idescat (padró municipal). -

A Bike Ride to Appreciate the Delta
Index 1. Terres de l’Ebre: destination cycling enthusiasts 03 2. Internationally recognised distinctive landscapes 04 3. The bicycle is the ideal means of transport to get to know Terres de l’Ebre 06 4. Receiving the visitor 08 5. Cycletourism in Terres de l’Ebre 10 6. Biking holiday deals 11 Ametlla de Mar Photograph yielded by Patronat de Turisme de la Diputació de Tarragona - Terres de l'Ebre 1. Terres de l’Ebre: destination cycling enthusiasts “You name it, here you have it” seems to be the case in these lands, and make them quite exceptional if not unique. Terres de l’Ebre is a mosaic, between the Mediterranean sea and the southern most part of the coastal Catalan ranges, whichever way you look at it, whether you are admiring its scenic beauty or its cultural and culinary life. Nor far removed from more frequented tourist resorts and built up areas, this relatively small area is criss-crossed by backroads, lanes and paths, ope- ning it up to the walker, cyclist and motorist alike. There really is a bit of everything in Terres de l’Ebre: ▶ First and foremost, the calm waters of the River ▶ Caves and World Heritage cave paintings bear wit- Ebro, that have shaped this land on the last stage of ness to the fact that Man has always lived here, cul- its journey to the sea. tivated the lands and lived well. ▶ Mile upon mile of largely unspoilt deserted bea- ▶ The episcopal and Renaissance city of Tortosa, an ches give way to a coastline carved out into lovely, example of once powerful, ancient cities.