Role of Leptin in Inflammation and Vice Versa
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Associations Between Serum Leptin Level and Bone Turnover in Kidney Transplant Recipients
Associations between Serum Leptin Level and Bone Turnover in Kidney Transplant Recipients ʈ ʈ ʈ Csaba P. Kovesdy,*† Miklos Z. Molnar,‡§ Maria E. Czira, Anna Rudas, Akos Ujszaszi, Laszlo Rosivall,‡ Miklos Szathmari,¶ Adrian Covic,** Andras Keszei,†† Gabriella Beko,‡‡ ʈ Peter Lakatos,¶ Janos Kosa,¶ and Istvan Mucsi §§ *Division of Nephrology, Salem Veterans Affairs Medical Center, Salem, Virginia; †Division of Nephrology, University of Virginia, Charlottesville, Virginia; ‡Institute of Pathophysiology, Semmelweis University, Budapest, Hungary; §Harold Simmons Center for Chronic Disease Research & Epidemiology, Los Angeles Biomedical Research Institute at ʈ Harbor-University of California–Los Angeles Medical Center, Torrance, California; Institute of Behavioral Sciences, Semmelweis University, Budapest, Hungary; ¶First Department of Internal Medicine, Semmelweis University, Budapest, Hungary; **University of Medicine Gr T Popa, Iasi, Romania; ††Department of Epidemiology, Maastricht University, Maastricht, Netherlands; ‡‡Central Laboratory, Semmelweis University, Budapest, Hungary; and §§Division of Nephrology, Department of Medicine, McGill University Health Center, Montreal, Quebec, Canada Background and objectives: Obesity is associated with increased parathyroid hormone (PTH) in the general population and in patients with chronic kidney disease (CKD). A direct effect of adipose tissue on bone turnover through leptin production has been suggested, but such an association has not been explored in kidney transplant recipients. Design, setting, participants, & measurements: This study examined associations of serum leptin with PTH and with biomarkers of bone turnover (serum beta crosslaps [CTX, a marker of bone resorption] and osteocalcin [OC, a marker of bone formation]) in 978 kidney transplant recipients. Associations were examined in multivariable regression models. Path analyses were used to determine if the association of leptin with bone turnover is independent of PTH. -

The Microbial Hypothesis: Contributions of Adenovirus Infection and Metabolic Endotoxaemia to the Pathogenesis of Obesity
Hindawi Publishing Corporation International Journal of Chronic Diseases Volume 2016, Article ID 7030795, 11 pages http://dx.doi.org/10.1155/2016/7030795 Review Article The Microbial Hypothesis: Contributions of Adenovirus Infection and Metabolic Endotoxaemia to the Pathogenesis of Obesity Amos Tambo, Mohsin H. K. Roshan, and Nikolai P. Pace Centre for Molecular Medicine and Biobanking, University of Malta, Msida, Malta Correspondence should be addressed to Amos Tambo; [email protected] Received 24 July 2016; Revised 10 October 2016; Accepted 25 October 2016 Academic Editor: Jochen G. Schneider Copyright © 2016 Amos Tambo et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. The global obesity epidemic, dubbed “globesity” by the World Health Organisation, is a pressing public health issue. The aetiology of obesity is multifactorial incorporating both genetic and environmental factors. Recently, epidemiological studies have observed an association between microbes and obesity. Obesity-promoting microbiome and resultant gut barrier disintegration have been implicated as key factors facilitating metabolic endotoxaemia. This is an influx of bacterial endotoxins into the systemic circulation, believed to underpin obesity pathogenesis. Adipocyte dysfunction and subsequent adipokine secretion characterised by low grade inflammation, were conventionally attributed to persistent hyperlipidaemia. They were thought of as pivotal in perpetuating obesity. It is now debated whether infection and endotoxaemia are also implicated in initiating and perpetuating low grade inflammation. The fact that obesity has a prevalence of over 600 million and serves as a risk factor for chronic diseases including cardiovascular disease and type 2 diabetes mellitus is testament to the importance of exploring the role of microbes in obesity pathobiology. -

Investigation of the Underlying Hub Genes and Molexular Pathogensis in Gastric Cancer by Integrated Bioinformatic Analyses
bioRxiv preprint doi: https://doi.org/10.1101/2020.12.20.423656; this version posted December 22, 2020. The copyright holder for this preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. Investigation of the underlying hub genes and molexular pathogensis in gastric cancer by integrated bioinformatic analyses Basavaraj Vastrad1, Chanabasayya Vastrad*2 1. Department of Biochemistry, Basaveshwar College of Pharmacy, Gadag, Karnataka 582103, India. 2. Biostatistics and Bioinformatics, Chanabasava Nilaya, Bharthinagar, Dharwad 580001, Karanataka, India. * Chanabasayya Vastrad [email protected] Ph: +919480073398 Chanabasava Nilaya, Bharthinagar, Dharwad 580001 , Karanataka, India bioRxiv preprint doi: https://doi.org/10.1101/2020.12.20.423656; this version posted December 22, 2020. The copyright holder for this preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. Abstract The high mortality rate of gastric cancer (GC) is in part due to the absence of initial disclosure of its biomarkers. The recognition of important genes associated in GC is therefore recommended to advance clinical prognosis, diagnosis and and treatment outcomes. The current investigation used the microarray dataset GSE113255 RNA seq data from the Gene Expression Omnibus database to diagnose differentially expressed genes (DEGs). Pathway and gene ontology enrichment analyses were performed, and a proteinprotein interaction network, modules, target genes - miRNA regulatory network and target genes - TF regulatory network were constructed and analyzed. Finally, validation of hub genes was performed. The 1008 DEGs identified consisted of 505 up regulated genes and 503 down regulated genes. -

Low Ambient Temperature Lowers Cholecystokinin and Leptin Plasma Concentrations in Adult Men Monika Pizon, Przemyslaw J
The Open Nutrition Journal, 2009, 3, 5-7 5 Open Access Low Ambient Temperature Lowers Cholecystokinin and Leptin Plasma Concentrations in Adult Men Monika Pizon, Przemyslaw J. Tomasik*, Krystyna Sztefko and Zdzislaw Szafran Department of Clinical Biochemistry, University Children`s Hospital, Krakow, Poland Abstract: Background: It is known that the low ambient temperature causes a considerable increase of appetite. The mechanisms underlying the changes of the amounts of the ingested food in relation to the environmental temperature has not been elucidated. The aim of this study was to investigate the effect of the short exposure to low ambient temperature on the plasma concentration of leptin and cholecystokinin. Methods: Sixteen healthy men, mean age 24.6 ± 3.5 years, BMI 22.3 ± 2.3 kg/m2, participated in the study. The concen- trations of plasma CCK and leptin were determined twice – before and after the 30 min. exposure to + 4 °C by using RIA kits. Results: The mean value of CCK concentration before the exposure to low ambient temperature was 1.1 pmol/l, and after the exposure 0.6 pmol/l (p<0.0005 in the paired t-test). The mean values of leptin before exposure (4.7 ± 1.54 μg/l) were also significantly lower than after the exposure (6.4 ± 1.7 μg/l; p<0.0005 in the paired t-test). However no significant cor- relation was found between CCK and leptin concentrations, both before and after exposure to low temperature. Conclusions: It has been known that a fall in the concentration of CCK elicits hunger and causes an increase in feeding activity. -

Leptin Replacement Reestablishes Brain Insulin Action in The
Diabetes Care 1 Sabine Frank-Podlech,1–3 Leptin Replacement Reestablishes Julia von Schnurbein,4 Ralf Veit,1–3 Martin Heni,2,3 Jurgen¨ Machann,2,5 Brain Insulin Action in the Jaana M. Heinze,2,3 Stephanie Kullmann,2,3 Jaida Manzoor,6 Hypothalamus in Congenital Saqib Mahmood,7 Hans-Ulrich Haring,¨ 2,3 fi Hubert Preissl,2,3,8,9 Martin Wabitsch,4 Leptin De ciency and Andreas Fritsche2,3 https://doi.org/10.2337/dc17-1867 OBJECTIVE Human obesity is associated with impaired central insulin signaling, and in very rare cases, severe obesity can be caused by congenital leptin deficiency. In such patients, leptin replacement results in substantial weight loss and improvement in peripheral 1 metabolism. Institute for Medical Psychology and Behaviou- ral Neurobiology, University of Tubingen,¨ Tubingen,¨ Germany RESEARCH DESIGN AND METHODS 2 Institute for Diabetes Research and Metabolic In a leptin-deficient patient, we investigated the impact of leptin substitution on Diseases of the Helmholtz Center Munich at the central insulin action, as quantified by changes in neuronal activity after intranasal University of Tubingen,¨ German Center for Dia- insulin application. This was assessed before and during the 1st year of metreleptin betes Research, Tubingen,¨ Germany 3Department of Internal Medicine IV, University substitution. Hospital, Tubingen,¨ Germany 4Division of Pediatric Endocrinology and Diabe- RESULTS tes, Department of Pediatrics and Adolescent After only 1 year, treatment with metreleptin reestablishes brain insulin sensitivity, Medicine, University of Ulm, Ulm, Germany 5 particularly in the hypothalamus and, to a lesser degree, in the prefrontal cortex. Section on Experimental Radiology, Depart- ment of Diagnostic and Interventional Radiol- Results are depicted in comparison with a control group. -

Metabolic Regulation of Fertility Through Presynaptic and Postsynaptic Signaling to Gonadotropin-Releasing Hormone Neurons
8578 • The Journal of Neuroscience, September 17, 2003 • 23(24):8578–8585 Behavioral/Systems/Cognitive Metabolic Regulation of Fertility through Presynaptic and Postsynaptic Signaling to Gonadotropin-Releasing Hormone Neurons Shannon D. Sullivan, R. Anthony DeFazio, and Suzanne M. Moenter 1Internal Medicine and Cell Biology, University of Virginia, Charlottesville, Virginia 22908 Gonadotropin-releasing hormone (GnRH) neurons form the final common pathway for the central regulation of reproduction and are inhibited by negative energy balance. In normal adults, these neurons maintain elevated intracellular chloride so that GABAA receptor activation is excitatory. We hypothesized that fasting alters homeostatic mechanisms to eliminate excitatory responses to GABA but rejected this hypothesis when brief, local GABA application elicited action currents in GnRH neurons from fed and fasted mice. This response was specific to GABAA receptors, because glycine elicited no response. We next found that fasting reduced the frequency of spontaneous GABAergic postsynaptic currents (PSCs) and that this was reversed by in vivo treatment with leptin during the fast. In the presence of tetrodotoxin to minimize presynaptic actions, leptin also potentiated the postsynaptic response of these cells to GABAA receptor activation. Postsynaptic effects of leptin on GABAergic miniature PSCs were eliminated by inhibiting JAK2/3 (Janus kinase), the tyrosine kinase through which leptin receptors signal. In all experiments, elimination of PSCs at ECl or by treatment with the GABAA receptor antagonist bicuculline confirmed that PSCs were specifically mediated by GABAA receptor chloride channels. These data dem- onstrate that fasting and leptin act presynaptically and postsynaptically to alter GABAergic drive to GnRH neurons, providing evidence for GABAergic communication of metabolic cues to GnRH neurons, and suggest the possibility for functional leptin receptors on GnRH neurons. -

High-Fat Diet-Induced Complement Activation Mediates Intestinal Inflammation and Neoplasia, Independent of Obesity Stephanie K
Published OnlineFirst August 17, 2016; DOI: 10.1158/1541-7786.MCR-16-0153 Metabolism Molecular Cancer Research High-Fat Diet-Induced Complement Activation Mediates Intestinal Inflammation and Neoplasia, Independent of Obesity Stephanie K. Doerner1, Edimara S. Reis2, Elaine S. Leung3, Justine S. Ko1, Jason D. Heaney4,5, Nathan A. Berger1,6, John D. Lambris2, and Joseph H. Nadeau1,3 Abstract Obesity and related metabolic disturbances are closely asso- and generation of C5a, which in turn induces the production of ciated with pathologies that represent a significant burden to proinflammatory cytokines and expression of proto-oncogenes. global health. Epidemiological and molecular evidence links Pharmacological and genetic targeting of the C5a receptor obesity and metabolic status with inflammation and increased reduced both inflammation and intestinal polyposis, suggest- risk of cancer. Here, using a mouse model of intestinal neo- ing the use of complement inhibitors for preventing diet- plasia and strains that are susceptible or resistant to diet- induced neoplasia. induced obesity, it is demonstrated that high-fat diet-induced inflammation, rather than obesity or metabolic status, is asso- Implications: This study characterizes the relations between diet ciated with increased intestinal neoplasia. The complement and metabolic conditions on risk for a common cancer and fragmentC5aactsasthetriggerforinflammation and intestinal identifies complement activation as a novel target for cancer tumorigenesis. High-fat diet induces complement activation prevention. Mol Cancer Res; 14(10); 953–65. Ó2016 AACR. Introduction Supporting a role for diet-induced obesity (DIO) in promoting inflammation and cancer, levels of IL6 and tumor burden are both Obesity is an increasingly important risk factor for many ob ob reduced following azoxymethane treatment of obese Lep / mice cancers (1, 2). -

Viral Infections and Interferons in the Development of Obesity
Tennessee State University Digital Scholarship @ Tennessee State University Agricultural and Environmental Sciences Department of Agricultural and Environmental Faculty Research Sciences 11-12-2019 Viral Infections and Interferons in the Development of Obesity Yun Tian Tennessee State University Jordan Jennings Tennessee State University Yuanying Gong Tennessee State University Yongming Sang Tennessee State University Follow this and additional works at: https://digitalscholarship.tnstate.edu/agricultural-and-environmental- sciences-faculty Part of the Diseases Commons, and the Immunology and Infectious Disease Commons Recommended Citation Tian, Y.; Jennings, J.; Gong, Y.; Sang, Y. Viral Infections and Interferons in the Development of Obesity. Biomolecules 2019, 9, 726. https://doi.org/10.3390/biom9110726 This Article is brought to you for free and open access by the Department of Agricultural and Environmental Sciences at Digital Scholarship @ Tennessee State University. It has been accepted for inclusion in Agricultural and Environmental Sciences Faculty Research by an authorized administrator of Digital Scholarship @ Tennessee State University. For more information, please contact [email protected]. biomolecules Review Viral Infections and Interferons in the Development of Obesity Yun Tian y, Jordan Jennings y, Yuanying Gong y and Yongming Sang * Department of Agricultural and Environmental Sciences, College of Agriculture, Tennessee State University, 3500 John A. Merritt Boulevard, Nashville, TN 37209, USA; [email protected] (Y.T.); [email protected] (J.J.); [email protected] (Y.G.) * Correspondence: [email protected]; Tel.: +1-615-963-5183 These authors contributed equally to this work. y Received: 14 October 2019; Accepted: 9 November 2019; Published: 12 November 2019 Abstract: Obesity is now a prevalent disease worldwide and has a multi-factorial etiology. -

Leptin As a Uremic Toxin Interferes with Neutrophil Chemotaxis
J Am Soc Nephrol 15: 2366–2372, 2004 Leptin as a Uremic Toxin Interferes with Neutrophil Chemotaxis LUCIANO OTTONELLO,* PAOLA GNERRE,* MARIA BERTOLOTTO,* MARINA MANCINI,* PATRIZIA DAPINO,* RODOLFO RUSSO,† GIACOMO GARIBOTTO,† TOMMASO BARRECA,* and FRANCO DALLEGRI* *Division of Internal Medicine and †Division of Nephrology, Department of Internal Medicine and Medical Specialties, University of Genoa Medical School, Genoa, Italy Abstract. Leptin is a pleiotropic molecule involved in energy Finally, the serum inhibitory activity can be effectively prevented homeostasis, hematopoiesis, inflammation, and immunity. Hypo- by immune depletion of leptin. The results also show, however, leptinemia characterizing starvation has been strictly related to that leptin by itself is endowed with chemotactic activity toward increased susceptibility to infection secondary to malnutrition. neutrophils. The two activities—inhibition of the cell response to Nevertheless, ESRD is characterized by high susceptibility to chemokines and stimulation of neutrophil migration—could be bacterial infection despite hyperleptinemia. Defects in neutrophils detected at similar concentrations. On the contrary, neutrophils 2ϩ play a crucial role in the infectious morbidity, and several uremic exposed to leptin did not display detectable [Ca ]i mobilization,  toxins that are capable of depressing neutrophil functions have oxidant production, or 2-integrin upregulation. The results dem- been identified. Only a few and contrasting reports about leptin onstrate that leptin is -
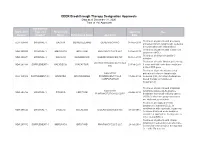
CDER Breakthrough Therapy Designation Approvals Data As of December 31, 2020 Total of 190 Approvals
CDER Breakthrough Therapy Designation Approvals Data as of December 31, 2020 Total of 190 Approvals Submission Application Type and Proprietary Approval Use Number Number Name Established Name Applicant Date Treatment of patients with previously BLA 125486 ORIGINAL-1 GAZYVA OBINUTUZUMAB GENENTECH INC 01-Nov-2013 untreated chronic lymphocytic leukemia in combination with chlorambucil Treatment of patients with mantle cell NDA 205552 ORIGINAL-1 IMBRUVICA IBRUTINIB PHARMACYCLICS LLC 13-Nov-2013 lymphoma (MCL) Treatment of chronic hepatitis C NDA 204671 ORIGINAL-1 SOVALDI SOFOSBUVIR GILEAD SCIENCES INC 06-Dec-2013 infection Treatment of cystic fibrosis patients age VERTEX PHARMACEUTICALS NDA 203188 SUPPLEMENT-4 KALYDECO IVACAFTOR 21-Feb-2014 6 years and older who have mutations INC in the CFTR gene Treatment of previously untreated NOVARTIS patients with chronic lymphocytic BLA 125326 SUPPLEMENT-60 ARZERRA OFATUMUMAB PHARMACEUTICALS 17-Apr-2014 leukemia (CLL) for whom fludarabine- CORPORATION based therapy is considered inappropriate Treatment of patients with anaplastic NOVARTIS lymphoma kinase (ALK)-positive NDA 205755 ORIGINAL-1 ZYKADIA CERITINIB 29-Apr-2014 PHARMACEUTICALS CORP metastatic non-small cell lung cancer (NSCLC) who have progressed on or are intolerant to crizotinib Treatment of relapsed chronic lymphocytic leukemia (CLL), in combination with rituximab, in patients NDA 206545 ORIGINAL-1 ZYDELIG IDELALISIB GILEAD SCIENCES INC 23-Jul-2014 for whom rituximab alone would be considered appropriate therapy due to other co-morbidities -

Review Article the Microbial Hypothesis: Contributions of Adenovirus Infection and Metabolic Endotoxaemia to the Pathogenesis of Obesity
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by OAR@UM Hindawi Publishing Corporation International Journal of Chronic Diseases Volume 2016, Article ID 7030795, 11 pages http://dx.doi.org/10.1155/2016/7030795 Review Article The Microbial Hypothesis: Contributions of Adenovirus Infection and Metabolic Endotoxaemia to the Pathogenesis of Obesity Amos Tambo, Mohsin H. K. Roshan, and Nikolai P. Pace Centre for Molecular Medicine and Biobanking, University of Malta, Msida, Malta Correspondence should be addressed to Amos Tambo; [email protected] Received 24 July 2016; Revised 10 October 2016; Accepted 25 October 2016 Academic Editor: Jochen G. Schneider Copyright © 2016 Amos Tambo et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. The global obesity epidemic, dubbed “globesity” by the World Health Organisation, is a pressing public health issue. The aetiology of obesity is multifactorial incorporating both genetic and environmental factors. Recently, epidemiological studies have observed an association between microbes and obesity. Obesity-promoting microbiome and resultant gut barrier disintegration have been implicated as key factors facilitating metabolic endotoxaemia. This is an influx of bacterial endotoxins into the systemic circulation, believed to underpin obesity pathogenesis. Adipocyte dysfunction and subsequent adipokine secretion characterised by low grade inflammation, were conventionally attributed to persistent hyperlipidaemia. They were thought of as pivotal in perpetuating obesity. It is now debated whether infection and endotoxaemia are also implicated in initiating and perpetuating low grade inflammation. -

A Microbiological Explanation for the Obesity Pandemic?
ADULT INFECTIOUS DISEASES NOTES A microbiological explanation for the obesity pandemic? Louis Valiquette MD MSC FRCPC1, Stéphanie Sirard MSc1, Kevin Laupland MD MSC FRCPC2,3 he prevalence of obesity is increasing worldwide. According to the two different and complementary mechanisms. They can extract TCanadian Health Measures Survey, in 2011, one in four Canadians energy from nondigestible polysaccharides and produce low-grade was obese (25% of women; 27% of men) (1). In addition to the imbal- inflammation (14). Because many Firmicutes are major butyrate pro- ance between energy intake and expenditure, sedentary lifestyle, a diet ducers, an abundance of bacteria from this phylum could be associated high in saturated fats and sugars, and genetic predisposition, many with an increase in genes encoding enzymes that enable the degrada- other factors may be involved in obesity. The presence of either sym- tion of complex polysaccharides and, in turn, increase the production biotic or pathogenic microorganisms may contribute to the develop- of monosaccharides and short-chain fatty acids (SCFAs) (15). Up to ment of obesity. With the progress of metagenomics and molecular 10% of the total energy extracted from food corresponds to SCFA techniques in recent years was born an interest in the microorganisms production (10). In a mouse model, the obese microbiome was found living on and inside us (2). More than 1014 microorganisms, which to be richer in enzymes involved in the digestion of complex polysac- represent up to 1150 different species and a total genome comprising charides. Consequently, higher concentrations of butyrate and acetate 150-fold more genes than the human genome, live in our gastrointes- were found in these mice cecum (16).