Prospecção Nutricional E Bioativa De Sementes De Dez Espécies Vegetais Da Caatinga
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Luan an Sau Hdcs
ư BỘ GIÁO DỤC VÀ ĐÀO TẠO BỘ NÔNG NGHIỆP VÀ PTNT VIỆN KHOA HỌC LÂM NGHIỆP VIỆT NAM NGUYỄN TH Ị LIỆU NGHIÊN CỨU CƠ SỞ KHOA HỌC VÀ KỸ THUẬT TRỒNG KEO LƯỠI LIỀM (Acacia crassicarpa A. Cunn. Ex. Benth) Ở VÙNG CÁT CHO MỤC ĐÍCH PHÒNG HỘ VÀ KINH TẾ TẠI TỈNH QUẢNG BÌNH, QUẢNG TRỊ VÀ THỪA THIÊN HUẾ LUẬN ÁN TIẾN SỸ LÂM NGHIỆP HÀ NỘI – 2017 BỘ GIÁO DỤC VÀ ĐÀO TẠO BỘ NÔNG NGHIỆP VÀ PTNT VIỆN KHOA HỌC LÂM NGHIỆP VIỆT NAM NGUYỄN THỊ LIỆU NGHIÊN CỨU CƠ SỞ KHOA HỌC VÀ KỸ THUẬT TRỒNG KEO LƯỠI LIỀM (Acacia crassicarpa A. Cunn. Ex. Benth) Ở VÙNG CÁT CHO MỤC ĐÍCH PHÒNG HỘ VÀ KINH TẾ TẠI TỈNH QUẢNG BÌNH, QUẢNG TRỊ VÀ THỪA THIÊN HUẾ Chuyên ngành: Lâm sinh Mã số: 62 62 02 05 Người hướng dẫn khoa học PGS. TS. Đặng Thái Dương HÀ NỘI – 2017 LỜI CAM ĐOAN Tôi xin cam đoan đây là công trình nghiên cứu khoa học của bản thân tôi, luận án được thực hiện trong thời gian từ năm 2013 đến 2017 dưới sự hướng dẫn của PGS.TS. Đặng Thái Dương. Các số liệu và kết quả nghiên cứ u trình bày trong luận án là trung thực. Nội dung của luận án có sử dụng một phần kết quả của đề tài nghiên cứu khoa học công nghệ cấp bộ "Điều tra tập đoàn cây trồng và xây dựng mô hình trồng rừng Keo lưỡi liềm (Acacia crassicarpa) trên cát nội đồng vùng Bắc Trung Bộ" do bản thân tác giả chủ trì. -

Adoption, Use and Perception of Australian Acacias Around the World
Diversity and Distributions, (Diversity Distrib.) (2011) 17, 822–836 S BIODIVERSITY Adoption, use and perception of PECIAL ISSUE RESEARCH Australian acacias around the world Christian A. Kull1*, Charlie M. Shackleton2, Peter J. Cunningham3, Catherine Ducatillon4, Jean-Marc Dufour-Dror5, Karen J. Esler6, James B. Friday7, Anto´nio C. Gouveia8, A. R. Griffin9, Elizabete Marchante8, Stephen J. :H Midgley10, Anı´bal Pauchard11, Haripriya Rangan1, David M. Richardson12, Tony Rinaudo13, Jacques Tassin14, Lauren S. Urgenson15, Graham P. von Maltitz16, Rafael D. Zenni17 and Matthew J. Zylstra6 UMAN - MEDIATED INTRODUCTIONS OF 1School of Geography and Environmental Science, ABSTRACT Monash University, Melbourne, Australia, 2Department of Environmental Science, Rhodes Aim To examine the different uses and perceptions of introduced Australian University, Grahamstown, South Africa, 3Sowing acacias (wattles; Acacia subgenus Phyllodineae) by rural households and Seeds of Change in the Sahel Project, SIM, communities. Maradi, Niger Republic, 4INRA, Jardin Thuret, Antibes, France, 5Environmental Policy Center, Location Eighteen landscape-scale case studies around the world, in Vietnam, Jerusalem Institute for Israel Studies, Jerusalem, India, Re´union, Madagascar, South Africa, Congo, Niger, Ethiopia, Israel, France, Israel, 6Department of Conservation Ecology and Portugal, Brazil, Chile, Dominican Republic and Hawai‘i. A Journal of Conservation Biogeography Entomology and Centre for Invasion Biology, Stellenbosch University, South Africa, 7University -

Proforma Reports for Applications to Clear Native Vegetation Under the Environmental Protection
Clearing Permit Decision Report 1. Application details 1.1. Permit application details Permit application No.: 6450/1 Permit type: Purpose Permit 1.2. Proponent details Proponent’s name: BHP Billiton Iron Ore Pty Ltd 1.3. Property details Property: Iron Ore (Goldsworthy-Nimingarra) Agreement Act 1972, Mining Lease 263SA (AM 70/263); Iron Ore (Goldsworthy-Nimingarra) Agreement Act 1972, Mineral Lease 251SA (AML 70/251); Iron Ore (Mount Goldsworthy) Agreement Act 1964, Mineral Lease 249SA (AML 70/249); Iron Ore (Mount Goldsworthy) Agreement Act 1964, Special Lease 3116/6935, Document J998594 L, Lot 42 on Deposited Plan 241586; Mining Leases 45/558, 45/573, 45/592, 45/1016, 45/1018; Exploration Licence 45/1072 Local Government Area: Shire of East PIlbara Colloquial name: Nimingarra to Yarrie Strategic Exploration Project 1.4. Application Clearing Area (ha) No. Trees Method of Clearing For the purpose of: 444.82 Mechanical Removal Mineral exploration, geotechnical investigations, hydrogeological drilling, access tracks, rehabilitation, and associated activities. 1.5. Decision on application Decision on Permit Application: Grant Decision Date: 26 March 2015 2. Site Information 2.1. Existing environment and information 2.1.1. Description of the native vegetation under application Vegetation Description Beard vegetation associations have been mapped for the whole of Western Australia. Three Beard vegetation associations have been mapped within the application area (GIS Database): 93: Hummock grasslands, shrub steppe; kanji over soft spinifex; 117: Hummock grasslands, grass steppe; soft spinifex; and 171: Hummock grasslands, low tree steppe; snappy gum over soft spinifex and Triodia brizioides. Eight flora and vegetation surveys have been undertaken over the application area by Onshore (2010, 2013, 2013a), Astron (2012, 2014), ENV (2008) and Ecologia (2005, 2005a). -
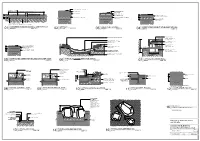
Port Hedland Landscape Guidelines Hardscape Elements Standard Drawing Details
PORT HEDLAND LANDSCAPE GUIDELINES HARDSCAPE ELEMENTS STANDARD DRAWING DETAILS STD01 - STD15 REV B 6th Feb 2018 1 of 2 varies @ A1 PORT HEDLAND LANDSCAPE GUIDELINES SOFTSCAPE ELEMENTS STANDARD DRAWING DETAILS STD16 - STD20 REV B 6th Feb 2018 2 of 2 varies @ A1 Appendix 3. Town of Port Hedland Preapproved Items ITEM DESCRIPTION PHOTO APPROVED MODEL/S 01 Compacted White Quartz Surfaces Cracker Dust 02 Compacted Red Scoria Surfaces 03 Concrete Grey or Red Tint Finishes 04 Concrete Shell Crete Finishes 05 Concrete Exposed Aggregate Finishes 06 Unit Pavers ‘Spinifex’ Paving 300x300x60mm 07 Edging Extruded Concrete – Bullnose edge 08 Edging 4mm Galvanised Steel 9 Compound 2.4m black Fencing galvanised palisade 10 Playground 2.4m black Fencing galvanised palisade- Flat top 11 Board Walks Replas Enduroplank - Grey 12 Boom Gates Leda Security Products – Model MSGF 13 Chicanes 14 Park Lighting - Greenfrog Systems Powered - Sentinel 15 Park Lighting - Greenfrog Systems Solar – Stealth 16 Bollards - Steel Galvanised Steel 17 Bollards - Flexi Pole Bollards – Recycled Charcoal Plastic Dimensions may vary and need Town approval 18 Seats Exteria Street and Park Outfitters – Parkway Seat Colour to be approved by the Town 19 Benches and Exteria Street and Tables Park Outfitters – Garden Table with Boardwalk Bench Colour to be approved by the Town 20 Boulder Seating Unquarried Sandstone Boulders 21 Litter Bin Post Exterior Street & Park Outfitters – Steel SHS Posts Hot dip galvanised finish 22 Dog Bag Woodlands AMS Dispensers Dog Bag Dispenser – Galvanised Steel. 23 Drinking Urban Fountains Fountains and Furniture – PF400 Pet Friendly Drinking Fountain. Steel RHS Mai Body. Hot Dip galvanised or powdercoat (colour to be approved by the Town) 24 Bicycle Racks Single Hoop Galvanised Bicycle Racks. -

Acacia Tumida Var. Tumida Occurrence Map
WATTLE Acacias of Australia Acacia tumida F.Muell. ex Benth. var. tumida Source: W orldW ideW attle ver. 2. Source: W orldW ideW attle ver. 2. Source: W orldW ideW attle ver. 2. Published at: w w w .w orldw idew attle.com Published at: w w w .w orldw idew attle.com Published at: w w w .w orldw idew attle.com B.R. Maslin B.R. Maslin Kym Brennan Source: W orldW ideW attle ver. 2. Published at: w w w .w orldw idew attle.com Kym Brennan Source: W orldW ideW attle ver. 2. Source: W orldW ideW attle ver. 2. Published at: w w w .w orldw idew attle.com Published at: w w w .w orldw idew attle.com B.R. Maslin See illustration. Source: W orldW ideW attle ver. 2. Source: W orldW ideW attle ver. 2. Published at: w w w .w orldw idew attle.com Published at: w w w .w orldw idew attle.com B.R. Maslin Kym Brennan Source: W orldW ideW attle ver. 2. Source: W orldW ideW attle ver. 2. Published at: w w w .w orldw idew attle.com Published at: w w w .w orldw idew attle.com See illustration. See illustration. Acacia tumida var. tumida occurrence map. O ccurrence map generated via Atlas of Living Australia (https://w w w .ala.org.au). Common Name Pindan Wattle, Sickle-leaf Wattle and more, fide M.W.McDonald, loc. cit. 158 Family Fabaceae Distribution Occurs in the Kimberley region of W.A. extending from Broome E to the Victoria R. -

Bardi Plants an Annotated List of Plants and Their Use
H.,c H'cst. /lust JIus lH8f), 12 (:J): :317-:359 BanE Plants: An Annotated List of Plants and Their Use by the Bardi Aborigines of Dampierland, in North-western Australia \!o\a Smith and .\rpad C. Kalotast Abstract This paper presents a descriptive list of the plants identified and used by the BarcE .\borigines of the Dampierland Peninsula, north~\q:stern Australia. It is not exhaust~ ive. The information is presented in two wavs. First is an alphabetical list of Bardi names including genera and species, use, collection number and references. Second is a list arranged alphabetically according to botanical genera and species, and including family and Bardi name. Previous ethnographic research in the region, vegetation communities and aspects of seasonality (I) and taxonomy arc des~ cribed in the Introduction. Introduction At the time of European colonisation of the south~west Kimberley in the mid nineteenth century, the Bardi Aborigines occupied the northern tip of the Dam pierland Peninsula. To their east lived the island-dwelling Djawi and to the south, the ~yulnyul. Traditionally, Bardi land ownership was based on identification with a particular named huru, translated as home, earth, ground or country. Forty-six bum have been identified (Robinson 1979: 189), and individually they were owned by members of a family tracing their ownership patrilineally, and known by the bum name. Collectively, the buru fall into four regions with names which are roughly equivalent to directions: South: Olonggong; North-west: Culargon; ~orth: Adiol and East: Baniol (Figure 1). These four directional terms bear a superficial resemblance to mainland subsection kinship patterns, in that people sometimes refer to themselves according to the direction in which their land lies, and indeed 'there are. -

Dampierland 2
Dampierland 2 Dampierland 2 (DL2 – Pindanland subregion) GORDON GRAHAM SEPTEMBER 2001 Subregional description and biodiversity • Adansonia gregorii (boab), Bauhinia cunninghamii and Grevillea striata (beefwood) grassy low open- values woodland. • Corymbia dampieri low open-woodland with Acacia spp. Shrubs and Triodia pungens (soft spinifex) and Description and area Triodia bitextura hummock grasses. • Eucalyptus brevifolia (snappy gum) low open- There are four basic components to the subregion. These woodland with Triodia spp. (spinifex) hummock comprise; grasses or sometimes a hummock grassland without • Quaternary sandplain overlying Jurassic and trees. Mesozoic sandstones with Pindan. There are • Acacia ancistrocarpa (Fitzroy wattle) and/or Acacia hummock grasslands on hills. eriopoda (Broome pindan wattle) and/or Acacia • Quaternary marine deposits on coastal plains, with monticola (Gawar) tall shrubland with Triodia mangal, samphire – Sporobolus spp. grasslands, intermedia (lobed spinifex) and Triodia pungens Melaleuca alsophila low forests, and Spinifex spp. – (soft spinifex) hummock grasses. Crotalaria spp. strand communities. • Grevillea refracta +/- Hakea lorea corkwood open- • Quaternary alluvial plains associated with the shrubland with Triodia pungens (soft spinifex) Permian and Mesozoic sediments of Fitzroy Trough hummock grasses. support tree savannahs of ribbon grass (Chrysopogon • Triodia pungens (soft spinifex) and/or Triodia spp.) – bluegrass (Dichanthium spp.) grasses with schinzii hummock grassland wooded with low trees scattered coolibah (Eucalyptus microtheca) - and Acacia spp. Shrubs. Bauhinia cunninghamii. There are riparian forests of river red gum (Eucalyptus camaldulensis) and Cadjeput (Melaleuca spp.) fringe drainages. Dominant land use (see Appendix B, key b) The climate is described as dry hot tropical and semi-arid with summer rainfall. The average annual rainfall is (ix) Grazing – Native pastures between 450 – 700 mm, slightly lower than the Fitzroy (xi) UCL and Crown reserves Trough subregion. -

Dear Metnkrs
ISeJ 10354638 ~mOFSOCIE;TIESm~~mAMS ACACIA~eWXlP~No.73- FEBRLlARY 1995 Dear Metnkrs, The 18th Biennial ASGAP Conference is at Ballarat on 23-30 September this year and as this is only a couple of hours drive away I hope to meet sane Acacia Study -up rnmcers there. I'm not sure yet which days or for how long I will be attenciing but I hope it will be for several days at least. Perhaps at sane stage Acacia Qoup members &nay be able to arrange an informal get together. If you ate attending perhaps you would like to help with the Study Group Display on the evening of Tuesday 26th. If so please let me !-am. It has been suggested that sane enlarged photos of Acacias be obtained and laminated for this and future displays. I have not got around to sarting out slides far this but if you have any very qccd slides or negatives of acacias growing in containers, cultivation (garden, park ar in xalsettings) wh.ich you muld allow the group to use please forward then far consideration (by the end of March), Forward and return postage will be paid by the -up. Close up photos of interesting aspects (e.g. seeds, leaves, flowers etc. 1 of acacias are also required, Slides to hand have enough good shots of plant farm etc. Bruce mlin (see below) has sent news of the future publication of the Acacia ~lmesof the "Flora of Australia" series. mis will give a treatment of all known species although I note new species of Eucalypts have been described since the publication of the volume on that genus. -

The Australian Centre for International Agricultural Research (ACIAR) Was Established in June 1982 by an Act of the Australian Parliament
The Australian Centre for International Agricultural Research (ACIAR) was established in June 1982 by an Act of the Australian Parliament. Its mandate is to help identify agricultural problems in developing countries and to commission collaborative research between Australian and developing country researchers in fields where Australia has a special research competence. Where trade names are used this does not constitute endorsement of nor discrimination against any product by the Centre. ACIAR PROCEEDINGS This series of publications includes the full proceedings of research workshops or symposia organised or supported by ACIAR. Numbers in this series are distrib uted internationally to selected individuals and scientific institutions. Previous numbers in the series are listed on the inside back cover. © Australian Centre for International Agricultural Research G.P.O. Box 1571, Canberra, A.C.T. 2601 Turnbull, John W. 1987. Australian acacias in developing countries: proceedings of an international workshop held at the Forestry Training Centre, Gympie, Qld., Australia, 4-7 August 1986. ACIAR Proceedings No. 16, 196 p. ISBN 0 949511 269 Typeset and laid out by Union Offset Co. Pty Ltd, Fyshwick, A.C.T. Printed by Brown Prior Anderson Pty Ltd, 5 Evans Street Burwood Victoria 3125 Australian Acacias in Developing Countries Proceedings of an international workshop held at the Forestry Training Centre, Gympie, Qld., Australia, 4-7 August 1986 Editor: John W. Turnbull Workshop Steering Committee: Douglas 1. Boland, CSIRO Division of Forest Research Alan G. Brown, CSIRO Division of Forest Research John W. Turnbull, ACIAR and NFTA Paul Ryan, Queensland Department of Forestry Cosponsors: Australian Centre for International Agricultural Research (ACIAR) Nitrogen Fixing Tree Association (NFTA) CSIRO Division of Forest Research Queensland Department of Forestry Contents Foreword J . -

Rangelands, Western Australia
Biodiversity Summary for NRM Regions Species List What is the summary for and where does it come from? This list has been produced by the Department of Sustainability, Environment, Water, Population and Communities (SEWPC) for the Natural Resource Management Spatial Information System. The list was produced using the AustralianAustralian Natural Natural Heritage Heritage Assessment Assessment Tool Tool (ANHAT), which analyses data from a range of plant and animal surveys and collections from across Australia to automatically generate a report for each NRM region. Data sources (Appendix 2) include national and state herbaria, museums, state governments, CSIRO, Birds Australia and a range of surveys conducted by or for DEWHA. For each family of plant and animal covered by ANHAT (Appendix 1), this document gives the number of species in the country and how many of them are found in the region. It also identifies species listed as Vulnerable, Critically Endangered, Endangered or Conservation Dependent under the EPBC Act. A biodiversity summary for this region is also available. For more information please see: www.environment.gov.au/heritage/anhat/index.html Limitations • ANHAT currently contains information on the distribution of over 30,000 Australian taxa. This includes all mammals, birds, reptiles, frogs and fish, 137 families of vascular plants (over 15,000 species) and a range of invertebrate groups. Groups notnot yet yet covered covered in inANHAT ANHAT are notnot included included in in the the list. list. • The data used come from authoritative sources, but they are not perfect. All species names have been confirmed as valid species names, but it is not possible to confirm all species locations. -

Biodiversity Summary: Rangelands, Western Australia
Biodiversity Summary for NRM Regions Guide to Users Background What is the summary for and where does it come from? This summary has been produced by the Department of Sustainability, Environment, Water, Population and Communities (SEWPC) for the Natural Resource Management Spatial Information System. It highlights important elements of the biodiversity of the region in two ways: • Listing species which may be significant for management because they are found only in the region, mainly in the region, or they have a conservation status such as endangered or vulnerable. • Comparing the region to other parts of Australia in terms of the composition and distribution of its species, to suggest components of its biodiversity which may be nationally significant. The summary was produced using the Australian Natural Natural Heritage Heritage Assessment Assessment Tool Tool (ANHAT), which analyses data from a range of plant and animal surveys and collections from across Australia to automatically generate a report for each NRM region. Data sources (Appendix 2) include national and state herbaria, museums, state governments, CSIRO, Birds Australia and a range of surveys conducted by or for DEWHA. Limitations • ANHAT currently contains information on the distribution of over 30,000 Australian taxa. This includes all mammals, birds, reptiles, frogs and fish, 137 families of vascular plants (over 15,000 species) and a range of invertebrate groups. The list of families covered in ANHAT is shown in Appendix 1. Groups notnot yet yet covered covered in inANHAT ANHAT are are not not included included in the in the summary. • The data used for this summary come from authoritative sources, but they are not perfect. -

Ecophysiological Responses of Three Species of Acacia
Central International Journal of Plant Biology & Research Bringing Excellence in Open Access Research Article *Corresponding author Abdulrahman A. Alzandi, Department of Biology, Al- Baha University, Saudi Arabia, Tel: 96-65057-78962; Email: Ecophysiological Responses of Submitted: 08 November 2018 Accepted: 21 November 2018 Three Species of Acacia Published: 26 November 2018 ISSN: 2333-6668 Abdulrahman A. Alzandi* Copyright Department of Biology, Al-Baha University, Saudi Arabia © 2018 Alzandi Abstract OPEN ACCESS Three species of Acacia (A .gerrardii, A. oerfota and A. origena) were evaluated Keywords for their ecophysiological behavior characteristics. Succulent, pigments contents, • Acacia minerals and some metabolic products were determined. The obtained data clearly • Succulence showed highest value of succulent (9.6%) in case of A. gerrardii. Concerning pigments • Pigments contents, current investigation revealed high contents of both chlorophyll a (1.72) mg/g • Minerals fresh weight and carotenoids (0.67) mg/g fresh weight in A. gerrardii as compared • Carbohydrates to the other two species. The obtained data recorded high content in chlorophyll b • Proteins (1.36) mg/g fresh weight in A. oerfota as compared to other species. With regard • Lipids to mineral composition, the obtained data show that the highest mineral percent was Calcium (3.8%) which detected in A. gerrardii followed by Potassium and Magnesium. Generally, A. gerrardii recorded highest percentages of all detected minerals. As regard to metabolic products, A. origena shows highest contents of total carbohydrates (46.47%) as well as total lipids (4.83%) as compared to the other species, while the highest crude protein value (34.77%) was recorded in A.